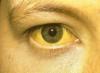
ДНХ-ийн хуулбар- энэ бол амьд организмын эсийг нөхөн үржих явцад эцэг эхийн ДНХ молекулыг хоёр дахин нэмэгдүүлэх үйл явц юм. Өөрөөр хэлбэл, хуулбарлах үйл явц нь эсийн хуваагдлаас өмнө явагддаг. Хуулбарлах нь транскрипци ба орчуулгатай адил юм матрицын процесс. Гинжин репликацийн явцад ДНХ молекулууд салж, тус бүр нь шинэ нэмэлт гинж нийлэгждэг загвар болж хувирдаг. Энэ тохиолдолд шинэ гинжний нуклеотидууд хослоно нэмэлтхуучин гинжин хэлхээний нуклеотидуудтай (A-тай T, G-тэй C). Үүний үр дүнд хоёр хэлхээтэй ДНХ-ийн хоёр охин молекул үүсдэг бөгөөд энэ нь үндсэн молекулаас ялгагдахгүй. ДНХ-ийн молекул бүр нь анхны эх молекулын нэг хэлхээ болон шинээр нийлэгжсэн нэг хэлхээнээс бүрдэнэ. Энэ хуулбарлах механизм гэж нэрлэгддэг хагас консерватив. Шинээр нийлэгжсэн хэлхээ бүр эсрэг параллельэцэг эх. Нэг хэлхээний синтез (тэргүүлэх нь) тасралтгүй явагддаг бол нөгөө нь (хоцрогдсон) импульс юм. Ийм механизмыг нэрлэдэг хагас тасралтгүй.
Хуулбарлах сэрээний бүтэц. Тэргүүлэх хэлхээ, хоцрогдсон хэлхээ, Оказаки хэлтэрхий. зургийг үзнэ үү.
ДНХ-ийн нийлэгжилтэнд оролцдог гол ферментүүд.
ДНХ полимеразын бүтцийн ерөнхий шинж чанарууд.
Тэд ижил зарчмаар ажилладаг: 3' төгсгөлд 1 нуклеотид нэмж ДНХ-ийн гинжийг уртасгадаг. Сонголт нь загвар ДНХ-ийн нэмэлт шаардлагаас хамаарна. Онцлогууд:
Хэд хэдэн бие даасан домэйн, cat. хамтдаа хүний баруун гартай төстэй. ДНХ нь гурван домайнаас үүссэн жижиг доголд холбогддог. Катализаторын төвийн үндэс нь далдуу модны домайн дахь консерватив амин хүчлийн сэдэл юм. "Хуруунууд" идэвхтэй төвд матрицыг зөв байрлуулна. "Эрхий хуруу" нь ферментийн гарц дээр ДНХ-ийг холбож, өндөр процессыг үүсгэдэг. Идэвхтэй төвд бүх гурван домэйны хамгийн чухал хамгаалагдсан бүсүүдийг нэгтгэж, тасралтгүй гадаргууг бүрдүүлдэг. Экзонуклеазын үйл ажиллагаа нь өөрийн катализаторын талбайтай бие даасан домэйнд байрладаг. N-терминал домэйн нь экзонуклеазын домайн руу ордог.
ДНХ полимераз I-ийн онцлог.
Гэмтсэн ДНХ-ийг нөхөн сэргээхэд оролцдог, мөн ДНХ-ийн хуулбарлахад туслах үүрэг гүйцэтгэдэг - загвар гинжтэй хосолсон гинжин хэлхээний 3' төгсгөлийг сунгаж, хоцрогдсон хэлхээний хэлтэрхийнүүдээр м/д цоорхойг дүүргэх, Оказаки уртасгах боломжийг олгодог. 3'-төгсгөлийн хэлтэрхийнүүд, нэгэн зэрэг РНХ-рибонуклеозидын үрийг устгаж, мууртай. Оказаки сегмент бүр эхэлнэ. ДНХ полимераз I нь давхар судалтай ДНХ-ийн завсарлагад нэг хэлхээний 3' төгсгөлийг сунгаж, мөн ижил завсарлагааны 5' төгсгөлөөс нуклеотидуудыг зайлуулж чаддаг (ник орчуулга), засварын системд чухал үүрэг гүйцэтгэдэг.
ДНХ-ийн полимер I бусад бүх зүйлийг давамгайлдаг. Энэ нь ойролцоогоор 103 кДа полипептид бөгөөд 2 хэсэгт хуваагдах боломжтой: C-терминал фрагмент 68 кДа, Кленовын хэсэг нь полимераз ба 3'->5" экзонуклеазын үйл ажиллагаатай; N-терминал фрагмент нь 35 кДа 5'->3" экзонуклеазын идэвхжил.
Холофермент, ДНХ полимераз III, реплисом.
Холоэнзим нь 4 төрлийн дэд цогцолборт хуваагддаг 10 уураг агуулсан 1900 кДа масстай цогцолбор юм.
α, ξ, θ. Катализаторын 2 хуулбарыг агуулна. α - ДНХ полимеразын идэвх, ξ - 3'-экзонуклеазын идэвх, θ - экзонуклеазыг өдөөдөг.
τ (tau) 2 дэд нэгжийг агуулдаг - катализаторын идэвхжил (α) бүхий хамгийн бага ферментийг холбоход үйлчилдэг.
Хавчаар (хавчаар) -ын 2 хуулбар - ДНХ-ийн загварт хамгийн бага ферментийг барих үүрэгтэй. Тус бүр нь β дэд нэгжийн гомодимерээс бүрдэнэ. Гол үүрэг нь хуулбарлах процесс дуусахаас өмнө матрицаас ферментийг салгах боломжийг багасгах явдал юм.
γ нь 5 уургийн бүлэг, муур. хавчаар ачигч үүсгэх - ДНХ матрицад хавчаар тавих төхөөрөмж. 2 δ, 1 γ, 1 ψ ба 1 χ - дэд нэгжээс бүрдэнэ.

Replisome - хагас консерватив хуулбарлах процессыг явуулдаг бактерийн хуулбарлах салаа дахь олон ферментийн цогцолбор; ДНХ полимераз болон бусад олон уураг агуулдаг.
эукариот ДНХ полимеразууд.
ДНХ полимераз α - шинэ хэлхээ ба хоцрогдсон хэлхээний нийлэгжилтийг эхлүүлдэг. β-дэд нэгж ба хоёр жижиг уурагтай холбогддог тул гинжийг дахин нэгтгэж чаддаг. 2 функц: праймер ба суналт = α-primase.
ДНХ полимераз δ - тэргүүлэх хэлхээг уртасгадаг
ДНХ полимераз ξ - хоцрогдсон хэлхээний нийлэгжилтэнд оролцдог
ДНХ-ийн лигазууд.
Репликаци, засвар, рекомбинацийн үед ДНХ-ийн гинжийг холбоход шаардлагатай. E. coli ба T4 фагийн ДНХ-ийн лигаза нь хоёр өөр дуплекс фрагментийн төгсгөл эсвэл шугаман эсвэл дугуй ДНХ-ийн гинжин хэлхээний тасархай төгсгөлийг холбох чадвартай дан пептидүүд юм. Иймээс шугаман болон дугуй хэлбэрийн дуплекс ДНХ молекулуудыг ДНХ-ийн лигазын тусламжтайгаар үүсгэж болно.
ДНХ геликазууд.
ATP гидролизийн энергийг ашиглан гинжийг задлах ажлыг гүйцэтгэдэг. Энэ нь хуулбарлах сэрээг хөдөлгөж, мушгиагүй гинжийг хуулбарлах цогцолборын нэг хэсэг болж ажилладаг. Хэд хэдэн Зеликасууд хурдыг нэмэгдүүлэхийн тулд хамтран ажиллах боломжтой.
SSB- уураг.
Нэг судалтай холбогч уургууд нь спиральыг тогтворгүй болгож, нэг судалтай бүсэд холбож, улмаар тогтворжуулдаг, i.e. нэг судалтай ДНХ-ийн хэсгийг засах.
ДНХ топоизомеразаIболонII, гираз.
ДНХ-ийг эргүүлээгүй үед молекул эргэлддэг - хоёрдогч болон гуравдагч бүтцийн өөрчлөлт. Эдгээр процессыг топоизомераза гэж нэрлэгддэг ферментийн бүлэг катализатор хийдэг. Тэд ДНХ-д нэг ба давхар хэлхээний завсарлага үүсгэдэг бөгөөд энэ нь нуклейн хүчлийн молекулыг эргүүлж, матриц болгох боломжийг олгодог. Үйлдлийн механизмын дагуу эхний (I) ба хоёр дахь (II) төрлийн топоизомераза нь ялгагдана.
I төрлийн топоизомераза (E. coli-д - swivelase) - ДНХ-ийн молекулд нэг хэлхээтэй тасалдлыг нэвтрүүлдэг, II төрлийн топоизомераза (E. coli-д - гираза) - давхар хэлхээтэй ДНХ-ийн тасалдал, ДНХ-ийн хэлхээг дамжуулдаг. завсарлага, дараа нь тэдгээрийн хөндлөн холбоос. Үүний зэрэгцээ, топизомераза нь үүргээ гүйцэтгэхдээ ДНХ-ийн молекултай холбоотой хэвээр байна. Эдгээр процессуудад топоизомераза нь фосфотирозин үүсэх замаар ДНХ фосфатын бүлэгт нуклеофилийн довтолгоо хийдэг тирозин үлдэгдлийг ашигладаг. Үүний үр дүнд ферментүүд нь ДНХ-ийн 5' эсвэл 3' төгсгөлтэй ковалент байдлаар холбогддог. Ийм ковалент холбоо үүсэх нь урвалын эцсийн үе шатанд нэг хэлхээний тасалдал дахь фосфодиэфирийн холбоог сэргээхэд эрчим хүчний зарцуулалтыг арилгадаг. I хэлбэрийн ДНХ топоизомераза нь нэг мономер уургийн молекул тутамд нэг каталитик тирозин үлдэгдэлтэй байдаг бол ДНХ топоизомераза II димерүүд нь дэд нэгж бүрт нэг катализаторын үлдэгдэл агуулдаг бөгөөд энэ нь ДНХ молекулд үе шаттайгаар давхар хэлхээний тасралт үүсгэдэг.
Топоизомераза нь нугасны үүрэг гүйцэтгэдэг боловч тэдгээрийн үйлдэл нь эсрэгээрээ байдаг. Топоизомераза I нь дугуй хэлбэрийн супер ороомогтой ДНХ-ийн нэг гинжийг тасалж, гинжийг задалж, супер ороомгийн тоог бууруулдаг. Топоизомераза II сул ороомоггүй битүү дугуй ДНХ-ийг супер ороомог болгон хувиргадаг.
Хуулбарлах үе шатууд: эхлэл, сунгалт, төгсгөл. ДНХ-ийн шинэ хэлхээ үүсэх эхлэл. Анхдагч. Примосом. Прокариотуудын ДНХ-ийн репликацийг дуусгавар болгох ба охин мушгиауудын ялгаа.
Шугаман геном дахь репликацийг зогсоох. Шугаман нээлттэй ДНХ-ийн фрагментийг хуулбарлах асуудал. Теломер ба теломерын давталт, теломерын гогцоо. Теломераза. Теломеразын механизм. Эукариот ДНХ-ийн хуулбарлах онцлог. эукариот хуулбарууд.
Бусад эсийн макромолекулуудын биосинтезийн нэгэн адил репликацийн процессыг нөхцөлт байдлаар хуваадаг. гурван үндсэн үе шат: эхлэл, сунгалт ба төгсгөл.
Прокариотуудын хуулбар
Санаачлага
Прокариот хромосомыг ихэвчлэн төлөөлдөг репликацийн нэг буюу хоёр гарал үүсэл бүхий нэг супер ороомог дугуй молекул.ДНХ-ийн хоёр хэлхээ бүр загвар болохын тулд шинэ хэлхээ нийлэгжүүлэхийн тулд ДНХ-ийн хэлхээ шулуун болж, бие биенээсээ холдох шаардлагатай. ДНХ-ийн гинж нь бүхэл бүтэн уртын дагуу биш, харин богино хэсэгт тайлагддаг болохыг тогтоожээ. Энд репликацын салаа үүсдэг - ДНХ-ийн хуулбарлах газар.
ДНХ-ийг эргүүлээгүй үед молекул эргэлддэг - хоёрдогч болон гуравдагч бүтцийн өөрчлөлт. Эдгээр процессыг ферментийн бүлэг гэж нэрлэдэг топоизомераза . Тэд ДНХ-д нэг ба давхар хэлхээний завсарлага үүсгэдэг бөгөөд энэ нь нуклейн хүчлийн молекулыг эргүүлж, матриц болгох боломжийг олгодог. Үйлдлийн механизмын дагуу эхний (I) ба хоёр дахь (II) төрлийн топоизомераза нь ялгагдана.
ДНХ-ийн эх молекулын репликаци эхэлдэг, репликацын гарал үүсэл (эсвэл гарал үүсэл, oriC) гэж нэрлэгддэг, мушгиагүй хэсэгт санаачлагч уургууд "суух". OriC дахь репликацийг эхлүүлэх нь DnaA, DnaB, DnaC, HU, gyrase, SSB гэсэн зургаан уураг агуулсан цогцолбор үүсэхээс эхэлдэг.
Уургууд эхлээд есөн нуклеотидын дараалалд холбогддог ДнаА , том дүүргэгчийг бүрдүүлдэг. Гарал үүслийн ДНХ нь түүнийг хүрээлж, ДНХ-ийн хэлхээ нь гурван 13-мер дарааллын бүсэд тусгаарлагддаг. Дараагийн шатанд DnaB (геликаз) ба DnaC уургууд холбогдож, 6 нм радиустай 480 кДа агрегатыг үүсгэдэг. Хеликаз/ DnaB ДНХ-ийн давхар хэлхээнд азотын суурийн хоорондох устөрөгчийн холбоог тасалж, түүний денатурацид хүргэдэг, өөрөөр хэлбэл. утаснуудын ялгаа.
ДНХ-ийн давхар мушгиа шулуун, денатурацийн үр дүнд Y хэлбэрийн репликацын салаа үүсдэг (Зураг 1). ДНХ полимеразууд нь энэ репликацийн салаа дээр охин ДНХ молекулуудыг нэгтгэдэг. ДНХ-ийн ийм хэсэг нь хуулбарлагдаагүй ДНХ-ийн бөмбөлөг эсвэл "нүд" шиг харагддаг. Хуулбарлах "нүд" нь хуулбарлах гарал үүслийн цэгүүд байрладаг газруудад үүсдэг. ДНХ-ийн хэлхээ тасрахад молекул нэлээд хөдөлгөөнтэй болдог. Үйлдлийн улмаас нэг хэлхээний бүтцэд гарч болзошгүй бүх эвдрэлийг оруулаагүй болно SSB уураг
(нэг судалтай ДНХ-тэй холбох уураг эсвэл спираль тогтворгүй болгох уураг) бөгөөд энэ нь ДНХ-ийн нэг хэлхээтэй холбогдож, хоорондоо наалдахаас сэргийлдэг. 
Энэ нь удамшлын молекул бөгөөд энэ чанарыг хэрэгжүүлэхийн тулд өөрийгөө яг хуулбарлах ёстой бөгөөд ингэснээр анхны ДНХ молекулд байгаа бүх мэдээллийг нуклеотидын тодорхой дараалал хэлбэрээр хадгалах ёстой. Үүнийг ДНХ-ийн репликац гэж нэрлэдэг биеийн аль ч эсийг хуваахаас өмнөх тусгай процессоор хангадаг.
ДНХ-ийн репликацийн мөн чанар нь тусгай фермент нь хоёр гинжин хэлхээний нуклеотидыг холбосон сул устөрөгчийн холбоог тасалдаг явдал юм. Үүний үр дүнд ДНХ-ийн хэлхээ тасарч, чөлөөт азотын суурь нь хэлхээ бүрээс "наалддаг" (репликацын салаа гэж нэрлэгддэг дүр төрх). Тусгай фермент, ДНХ полимераз нь чөлөөт ДНХ-ийн гинжин хэлхээний дагуу 5-аас 3-т (тэргүүлэх хэлхээ) хөдөлж, эсэд байнга нийлэгждэг чөлөөт нуклеотидуудыг шинээр нийлэгжсэн 3" төгсгөлд нэгтгэхэд тусалдаг. ДНХ-ийн хэлхээ ДНХ-ийн хоёр дахь хэлхээнд (хоцрогдсон хэлхээ) 1000-2000 нуклеотид (Оказаки хэлтэрхий) -ээс бүрдсэн жижиг сегмент хэлбэрээр шинэ ДНХ үүсдэг.
Энэ хэлхээний ДНХ-ийн хэсгүүдийн хуулбарыг эхлүүлэхийн тулд тусгай фермент - РНХ полимераз (примаза) -ийг ашигладаг үрийн хувьд богино РНХ фрагментуудын нийлэгжилтийг (РНХ-ийн онцлог шинж чанаруудыг доор авч үзэх болно) хийх шаардлагатай. Дараа нь РНХ праймеруудыг зайлуулж, ДНХ полимераз I ашиглан үүссэн цоорхойд ДНХ-ийг оруулна. Тиймээс ДНХ-ийн хэлхээ бүрийг нэмэлт хэлхээ үүсгэх загвар эсвэл загвар болгон ашигладаг бөгөөд ДНХ-ийн репликац нь хагас консерватив (жишээ нь нэг хэлхээ) юм. шинэ ДНХ молекул дахь "хуучин", хоёр дахь нь шинэ).
Эсийн тэргүүлэх болон хоцрогдсон хэлхээг хуулбарлахад янз бүрийн ферментүүдийг ашигладаг. Репликацийн үр дүнд хоёр шинэ туйлын ижил ДНХ молекул үүсдэг бөгөөд тэдгээр нь репликаци эхлэхээс өмнө анхны ДНХ молекултай ижил байдаг (ДНХ-ийн хуулбарлах үйл явцыг 3.5-р зурагт илүү дэлгэрэнгүй харуулав). ДНХ полимераз нь бусад ферментийн нэгэн адил чөлөөт ДНХ-ийн гинжин хэлхээнд нэмэлт нуклеотид нэмэх процессыг ихээхэн хурдасгадаг боловч аденин нь тимин, цитозины гуанинтай химийн хамаарал нь маш өндөр тул тэдгээр нь бие биетэйгээ нийлдэг. энгийн урвалын хольцод ДНХ полимераз байхгүй.
Энэ молекулын суурийн нэмэлт дээр үндэслэсэн ДНХ молекулын яг хоёр дахин нэмэгдэх үзэгдэл нь удамшлын молекул үндэс юм гэж зарим талаар хялбаршуулсан гэж хэлж болно. Хүний ДНХ-ийн репликацийн хурд харьцангуй удаан бөгөөд хэрэв репликаци нь нэг цэгээс эхэлсэн бол хүний аливаа хромосомын ДНХ-ийг хуулбарлахад долоо хоног шаардагдана. Үнэн хэрэгтээ аливаа хромосомын ДНХ-ийн молекулд хүний хромосом бүр зөвхөн нэг ДНХ молекул агуулдаг тул репликацын эхлэл (репликон) олон газар байдаг. Репликон бүрээс зэргэлдээх репликонууд нэгдэх хүртэл хоёр чиглэлд хуулбарлагдана. Тиймээс хромосом бүрт ДНХ-ийн хуулбар харьцангуй хурдан явагддаг.
Репликаци (лат. replicatio - шинэчлэх) - эх ДНХ молекулын матриц дээр охин ДНХ молекулыг нэгтгэх үйл явц. Эх эсийн дараагийн хуваагдлын явцад охин эс бүр анхны эх эсийн ДНХ-тэй ижил ДНХ молекулын нэг хувийг хүлээн авдаг. Энэ үйл явц нь генетикийн мэдээллийг үеэс үед үнэн зөв дамжуулах боломжийг олгодог. ДНХ-ийн репликацийг реплисом (англи хэлээр replisome) гэж нэрлэдэг 15-20 өөр уурагаас бүрдэх ферментийн цогц цогцолбор гүйцэтгэдэг.
ДНХ-ийн хуулбар нь эсийн хуваагдлын гол үйл явдал юм. Хуваагдах үед ДНХ бүрэн хуулбарлагдах бөгөөд зөвхөн нэг удаа байх нь чухал юм. Үүнийг ДНХ-ийн хуулбарлах зохицуулалтын тодорхой механизмаар хангадаг.
Хуулбарлах нь дотор явагддаг гурван үе шат:
1. Хуулбарлах эхлэл
2. Сунгах
3. Хуулбарлах ажиллагааг зогсоох.
Хуулбарлах ажлыг голчлон эхлүүлэх үе шатанд зохицуулдаг. Үүнийг хэрэгжүүлэхэд тун хялбар, учир нь репликаци нь ДНХ-ийн аль ч сегментээс биш, харин репликацийг эхлүүлэх газар гэж нэрлэгддэг нарийн тодорхойлогдсон хэсгээс эхэлж болно. Геномд ийм сайтууд зөвхөн нэг эсвэл олон байж болно. Хуулбарлах эхлэлийн талбай гэсэн ойлголт нь репликон гэсэн ойлголттой нягт холбоотой. Репликон гэдэг нь репликацийн эхлэлийн талбайг агуулсан ДНХ-ийн суналт бөгөөд энэ сайтаас ДНХ-ийн нийлэгжилт эхэлсний дараа хуулбарлагддаг. Бактерийн геном нь дүрмээр бол нэг репликон бөгөөд энэ нь бүх геномын репликаци нь зөвхөн нэг репликацийг эхлүүлсний үр дүн гэсэн үг юм.
Эукариот геномууд (түүнчлэн тэдгээрийн бие даасан хромосомууд) олон тооны бие даасан репликонуудаас бүрддэг бөгөөд энэ нь бие даасан хромосомын хуулбарлах нийт хугацааг мэдэгдэхүйц бууруулдаг. Эсийн хуваагдлын мөчлөг бүрт сайт тус бүр дээр репликацын эхлэлийн тоог хянадаг молекулын механизмыг хуулбарлах тоон хяналт гэж нэрлэдэг. Хромосомын ДНХ-ээс гадна бактерийн эсүүд ихэвчлэн бие даасан хуулбар болох плазмид агуулдаг. Плазмидууд нь хуулбарын дугаарыг хянах өөрийн гэсэн механизмтай байдаг: тэд нэг эсийн мөчлөгт плазмидын зөвхөн нэг хувь буюу мянга мянган хуулбарыг нийлэгжүүлж чаддаг.
Репликаци нь ДНХ-ийн давхар мушгиа задарч, ДНХ-ийн шууд хуулбарлах цэг болох репликацын салаа үүсгэснээр репликацийг эхлүүлсэн газраас эхэлдэг. Сайт бүр хуулбарлах нь нэг чиглэлтэй эсвэл хоёр чиглэлтэй эсэхээс хамааран нэг эсвэл хоёр репликацын салаа үүсгэж болно. Хоёр чиглэлтэй хуулбарлах нь илүү түгээмэл байдаг. Репликаци эхэлснээс хойш хэсэг хугацааны дараа электрон микроскопоор хуулбарлах нүдийг ажиглаж болно - ДНХ аль хэдийн хуулбарлагдсан хромосомын бүс, хуулбарлагдаагүй ДНХ-ийн илүү өргөтгөсөн хэсгүүдээр хүрээлэгдсэн.
Репликацийн салаа хэсэгт ДНХ нь том уургийн цогцолборыг (реплисом) хуулбарладаг бөгөөд гол фермент нь ДНХ полимераз юм. Репликацийн салаа нь прокариотуудад минутанд 100,000 суурь хос, эукариотуудад 500-5000 орчим суурь хос хурдтайгаар хөдөлдөг.
Молекулын хуулбарлах механизм:
Ферментүүд (геликаз, топоизомераза) болон ДНХ-г холбодог уурагууд нь ДНХ-ийг задалж, матрицыг шингэрүүлсэн төлөвт байлгаж, ДНХ-ийн молекулыг эргүүлдэг. Репликацийн зөв байдал нь нэмэлт суурь хосуудын яг таарч, алдааг таньж, засах чадвартай ДНХ полимеразын идэвхжилээр баталгааждаг. Эукариотуудын репликацийг хэд хэдэн өөр ДНХ полимеразууд гүйцэтгэдэг (прокариотуудын ДНХ-ийн репликациас ялгаатай).
ДНХ полимераз I нь хоцрогдсон хэлхээнд үйлчилж, РНХ праймеруудыг устгаж, цэвэршүүлсэн ДНХ-ийн хэсгүүдийг урьдчилан хуулбарлана. ДНХ полимераз III нь хоцрогдсон хэлхээний нийлэгжилтийн явцад тэргүүлэх ДНХ-ийн хэлхээ болон Оказаки фрагментуудыг нэгтгэдэг ДНХ-ийн хуулбарлах гол фермент юм (Оказаки хэлтэрхийнүүд нь ДНХ-ийн репликацийн явцад хоцрогдсон хэлхээ дээр үүсдэг харьцангуй богино ДНХ фрагментүүд юм). Дараа нь нийлэгжсэн молекулуудыг ДНХ-ийн хэт ороомог, цаашдын нягтралын зарчмын дагуу мушгина. Синтез нь эрчим хүч их шаарддаг.
ДНХ-ийн молекулын гинж нь хуваагдаж, репликацын салаа үүсгэдэг бөгөөд тэдгээр нь тус бүр нь шинэ нэмэлт гинж нийлэгждэг загвар болж хувирдаг. Үүний үр дүнд үндсэн молекултай ижил хоёр хэлхээтэй ДНХ-ийн шинэ молекулууд үүсдэг.
хуулбарлах- ДНХ-ээс ДНХ руу мэдээлэл дамжуулах, ДНХ-ийн өөрөө хуулбарлах (ДНХ биосинтез).
ДНХ-ийн молекул нь хоёр хэлхээнээс тогтдог хоёр дахин нэмэгддэгэсийн хуваагдлын үед. ДНХ хоёр дахин нэмэгддэгутас нэхэх үед утас бүрийг дуусгаж болно гэсэн баримт дээр тулгуурладаг нэмэлт хуулбар, ингэснээр ДНХ молекулын хоёр хэлхээг олж авч, эхийг нь хуулна.
Хуулбарлахад шаардлагатай нөхцөлүүд: 1.) Матриц- ДНХ-ийн хэлхээ. Утас хуваах гэж нэрлэдэг хуулбарлах сэрээ. Энэ нь ДНХ молекул дотор үүсч болно. Тэд янз бүрийн чиглэлд хөдөлж, үүсдэг хуулбарлах нүд. Эукариотуудын ДНХ молекулд хэд хэдэн ийм нүд байдаг бөгөөд тус бүр нь хоёр салаатай байдаг. 2.) субстрат. Хуванцар материал нь дезоксинуклеотид трифосфатууд: dATP, dGTP, dCTP, dTTP. Дараа нь тэд задардаг дезоксинуклеотид монофосфатууд, энерги ялгарах органик бус фосфатын хоёр молекул, i.e. тэд хоёулаа эх сурвалж мөн эрчим хүч, ба хуванцар материал. 3.) Ионууд магни. 4.) Хуулбарлах ферментийн цогцолбор. а) ДНХ уурагуудыг задалдаг: - ДНХ-А(утас хоорондын зөрүүг үүсгэдэг); - мушгиа(ДНХ-ийн хэлхээг таслах) - топоизомераза 1болон 2 (спираль дээгүүр тайвшрах). салгах (3,5")-фосфодиэфирийн холбоо. Прокариотуудын топоизомераза 2 гэж нэрлэгддэг гираза. б) ДНХ-ийн хэлхээг холбохоос сэргийлдэг уураг ( SSB уураг). онд) ДНХ полимераз(фосфодиэфирийн холбоо үүсэхийг катализатор). ДНХ полимераз нь аль хэдийн байгаа хэлхээг уртасгадаг боловч хоёр чөлөөт нуклеотидыг холбож чадахгүй. G) Примаза(нийлэгжүүлэх "үр" үүсэхийг хурдасгадаг). Энэ нь нэг нуклеотидыг холбодог РНХ полимеразын бүтцэд байдаг. д) ДНХ лигаз. 5.) Праймерууд- хуулбарлах "үр". Энэ бол товчхон хэсэг юм рибонуклеотид трифосфатууд(2 - 10). Праймер үүсэх нь катализатор юм анхдагч.
Хуулбарлах алхамууд: 1.) Санаачлага(хуулбарлах салаа үүсэх); 2.) Сунгах(шинэ утаснуудын синтез); 3.) Праймерыг хасах; 4.) Төгсгөл(хоёр хүүхдийн гинжин хэлхээний синтезийг дуусгах).
Хуулбарлах эхлэл:- дохионы уургийн молекулуудыг зохицуулах; өсөлтийн хүчин зүйлүүд;- хангах ферментүүдболон тусгай уураг.
Шаардлагатай ферментүүд: ДНХ топоизомеразаДНХ-ийн супер ороомогуудыг задалдаг ферментүүд. ДНХ-ийн геликаз- давхар судалтай ДНХ молекул дахь устөрөгчийн холбоог тасалдаг. Үүний үр дүнд А хуулбарлах сэрээ (хуулбарлах нүд).

Нэг хэлхээтэй ДНХ-ийн хэлхээтэй холбогддог уураг нь нэг хэлхээтэй ДНХ-тэй холбогдож, тэдгээрийн нэмэлт нэгдэлд саад болдог.
хуулбарлах суналт.Синтезийн субстратууд нь дезоксинуклеозид трифосфатууд, барилгын материал, эрчим хүчний эх үүсвэрийн үүрэг гүйцэтгэдэг.
Шаардлагатай ферментүүд: ДНХ-ийн примаз, ДНХ полимеразын РНХ праймерын богино молекулуудын нийлэгжилтийг хурдасгадаг. ДНХ полимеразөсөн нэмэгдэж буй "шинэ" гинжин хэлхээнд нэмэлт нуклеотидын "хуучин", өөрөөр хэлбэл загвар гинжийг оруулахыг баталгаажуулдаг.
ДНХ-ийн шинэ хэлхээний нийлэгжилт нь зөвхөн чиглэлд л үргэлжилж болно 5' төгсгөлөөс 3' төгсгөл хүртэл. Нэг хэлхээ дээр ДНХ тасралтгүй нийлэгждэг "тэргүүлэх" хэлхээ, нөгөө талд нь богино хэлтэрхийнүүд үүсдэг - "хоцрогдсон" гинж (Оказакигийн хэсгүүд).
Праймеруудыг арилгасны дараа ДНХ лигазбогино Оказаки хэлтэрхийг хооронд нь оёдог ( дуусгавар болгох).
Мэдээлэл дамжуулж байна матрицын арга. хагас консервативДНХ-ийн хуулбарлах механизм.
| Хоцрогдсон хэлхээний синтез |
| 3’ |
| 3’ |
| 5’ |
| 5’ |


ДНХ бол удамшлын мэдээллийн найдвартай нөөц юм. Гэхдээ үүнийг зөвхөн аюулгүй байлгахаас гадна үр удамд нь үлдээх ёстой. Энэ зүйлийн оршин тогтнох эсэх нь үүнээс хамаарна. Эцсийн эцэст эцэг эхчүүд хувьслын явцад олж авсан бүх зүйлээ хүүхдүүддээ дамжуулах ёстой. Үүнд: мөчний тооноос эхлээд нүдний өнгө хүртэл бүх зүйл бичигдсэн байдаг. Мэдээжийн хэрэг, бичил биетүүд энэ мэдээлэлээс хамаагүй бага байдаг, гэхдээ энэ нь бас дамжих ёстой. Үүнийг хийхийн тулд эс хуваагдана. Удамшлын мэдээлэл хоёр охин эсэд очихын тулд үүнийг хоёр дахин нэмэгдүүлэх шаардлагатай бөгөөд энэ үйл явцыг "ДНХ-ийн хуулбар" гэж нэрлэдэг. Энэ нь аль нь ч хамаагүй эсийн хуваагдахаас өмнө тохиолддог. Энэ нь үржихээр шийдсэн нян байж болно. Эсвэл зүссэн хэсэгт арьсны шинэ ургалт байж болно. Эсийн хуваагдал эхлэхээс өмнө дезоксирибонуклеины хүчлийн давхардлын процессыг тодорхой зохицуулж, дуусгах ёстой.
Хоёр дахин нэмэгдэх нь хаана тохиолддог вэ?
ДНХ-ийн хуулбар шууд цөмд (эукариотуудад) эсвэл цитоплазмд (прокариотуудад) явагддаг. Нуклейн хүчил нь нуклеотидуудаас бүрддэг - аденин, тимин, цитозин, гуанин. Молекулын хоёр гинж нь бие биенээ нөхөх зарчмын дагуу бүтээгдсэн байдаг: нэг гинж дэх аденин нь тиминтэй, гуанин нь цитозинтэй тохирдог. Молекулын хоёр дахин нэмэгдэл нь охин спиральд нэмэлт байх зарчим хадгалагдахаар явагдах ёстой.

Хуулбарлах эхлэл - эхлэл
Дезоксирибонуклеины хүчил нь хоёр судалтай спираль юм. ДНХ-ийн репликаци нь эцэг эхийн хэлхээ бүрийн дагуу хүүхдийн хэлхээг барьж дуусгаснаар явагддаг. Энэ нийлэгжилтийг боломжтой болгохын тулд спиральуудыг "тайлж", гинж нь бие биенээсээ тусгаарлагдсан байх ёстой. Энэ үүргийг геликаз гүйцэтгэдэг - энэ нь дезоксирибонуклеины хүчлийн мушгиа тайлж, өндөр хурдтайгаар эргэлддэг. ДНХ-ийн давхардлын эхлэл нь ямар ч газраас эхэлж болохгүй, ийм нарийн төвөгтэй үйл явц нь молекулын тодорхой хэсгийг шаарддаг - репликацийг эхлүүлэх газрыг шаарддаг. Давхардлын эхлэлийн цэгийг тодорхойлж, геликаз нь мушгиа тайлах ажлыг эхлүүлсний дараа ДНХ-ийн хэлхээ нь хажуу тийшээ хуваагдан репликацын салаа үүсгэдэг. ДНХ полимеразууд тэдгээрийн дээр суудаг. Тэд бол хүүхдийн гинжийг нэгтгэх болно.

Сунгах

Дезоксирибонуклеины хүчлийн нэг молекулд 5-аас 50 хүртэлх репликацын салаа үүсч болно. Охин гинжний синтез нь молекулын хэд хэдэн бүсэд нэгэн зэрэг явагддаг. Гэвч нэмэлт нуклеотидын бүтээн байгуулалтыг дуусгана гэдэг амаргүй. Нуклейн хүчлийн гинж нь бие биенээсээ эсрэг байдаг. Эцэг эхийн гинжин хэлхээний өөр өөр чиглэл нь хоёр дахин нэмэгдэхэд нөлөөлдөг бөгөөд энэ нь ДНХ-ийн хуулбарлах нарийн төвөгтэй механизмд хүргэсэн. Нэг гинжийг хүүхдийн гинж тасралтгүй дүүргэж, удирдагч гэж нэрлэдэг. Энэ нь зөв, учир нь полимераз нь чөлөөт нуклеотидыг өмнөх 3'-OH төгсгөлд хавсаргахад маш тохиромжтой. Ийм синтез нь хоёр дахь гинжин хэлхээний процессоос ялгаатай нь тасралтгүй үргэлжилдэг.
Хоцрогдсон гинж, О'косаксын хэлтэрхий
Чөлөөт нуклеотидыг холбох боломжгүй чөлөөт 5' төгсгөл байдаг тул нөгөө гинжин хэлхээнд хүндрэл гардаг. Дараа нь ДНХ полимераз нөгөө талд үйлчилнэ. Хүүхдийн хэлхээг дуусгахын тулд эцэг эхийн хэлхээнд нэмэлт праймерыг бий болгодог. Энэ нь өөрөө хуулбарлах салаа дээр үүсдэг. Жижиг хэсгүүдийн нийлэгжилт үүнээс эхэлдэг, гэхдээ аль хэдийн "зөв" зам дагуу - нуклеотидын нэмэлт нь 3' төгсгөлд тохиолддог. Тиймээс, хоёр дахь охин мушгиа дахь гинжин хэлхээг дуусгах нь тасалдалтай тохиолддог бөгөөд хуулбарлах сэрээний хөдөлгөөний эсрэг чиглэлтэй байдаг. Эдгээр хэсгүүдийг О'коссак хэлтэрхий гэж нэрлэдэг бөгөөд урт нь 100 орчим нуклеотид юм. Фрагментийг өмнөх бэлэн хэсэг болгон хийж дууссаны дараа праймеруудыг тусгай ферментээр хайчилж, зүссэн хэсгийг алга болсон нуклеотидуудаар дүүргэнэ.
Төгсгөл
Хоёр гинж нь хүүхдүүдээ дуусгахад хоёр дахин нэмэгдэж дуусч, О'коссакийн бүх хэлтэрхийнүүд хоорондоо оёж байна. Эукариотуудад ДНХ-ийн репликаци нь репликацын салаа бие биетэйгээ уулзах үед дуусдаг. Мөн прокариотуудад энэ молекул нь дугуй хэлбэртэй бөгөөд түүний хоёр дахин нэмэгдэх процесс нь эхлээд гинжийг таслахгүйгээр явагддаг. Бүх дезоксирибонуклеины хүчил нь нэг том хуулбар юм. Мөн хувилах салаа нь цагирагийн эсрэг талд нийлснээр хоёр дахин нэмэгддэг. Репликаци дууссаны дараа үндсэн дезоксирибонуклеины хүчлийн хоёр хэлхээ нь холбосон байх ёстой бөгөөд үүний дараа хоёр молекулыг мушгиж супер ороомог үүсгэдэг. Дараа нь ДНХ-ийн молекулын аль алиных нь метилжилт нь -GATC- сайт дахь аденин дээр явагддаг. Энэ нь гинжийг салгахгүй бөгөөд тэдгээрийн нэмэлт байдалд саад болохгүй. Энэ нь молекулуудыг хромосом болгон нугалах, мөн генийн уншилтыг зохицуулахад шаардлагатай.
Хуулбарлах хурд ба нарийвчлал

ДНХ-ийн давхардлын хоёр дахь үе шат (суналт) нь секундэд 700 орчим нуклеотидын хурдаар явагддаг. Хэрэв бид нуклейн хүчлийн нэг эргэлтэнд 10 хос мономер байдгийг санаж байвал молекул нь "муйрах" үед секундэд 70 эргэлтийн давтамжтайгаар эргэлддэг. Харьцуулбал: компьютерийн системийн нэгж дэх хөргөгчийн эргэлтийн хурд нь секундэд ойролцоогоор 500 эргэлт юм. Гэхдээ өндөр хурдтай байсан ч ДНХ полимераз бараг хэзээ ч буруу байдаггүй. Эцсийн эцэст тэр зүгээр л нэмэлт нуклеотидыг сонгодог. Гэвч тэр алдаа гаргасан ч ДНХ полимераз түүнийг таньж, нэг алхам ухарч, буруу мономерыг урж, зөвөөр сольдог. ДНХ-ийн хуулбарлах механизм нь маш нарийн төвөгтэй боловч бид гол санааг гаргаж чадсан. Энэ нь бичил биетэн болон олон эст организмын хувьд түүний ач холбогдлыг ойлгох нь чухал юм.
ДНХ-ийн хуулбарнь эс хуваагдахаас өмнө хоёр дахин нэмэгдэх үйл явц юм. Заримдаа "ДНХ-ийн хуулбар" гэж нэрлэдэг. Давхардах нь эсийн мөчлөгийн интерфазын S үе шатанд тохиолддог.
Зэрлэг ан амьтдын генетикийн материалыг өөрөө хуулбарлах нь мэдээжийн хэрэг. Зөвхөн ийм байдлаар л эсийн хуваагдлын явцад үүссэн охин эсүүд анхных шигээ их хэмжээний ДНХ агуулж чадна. Репликацийн ачаар бүтэц, бодисын солилцооны генетикийн програмчлагдсан бүх шинж чанарууд хэд хэдэн үе дамжин дамждаг.
Эсийн хуваагдлын явцад ижил төрлийн хос ДНХ-ийн молекул бүр охин эс рүүгээ очдог. Энэ нь удамшлын мэдээллийг үнэн зөв дамжуулах боломжийг олгодог.
ДНХ-ийн синтезийн явцад энерги зарцуулагддаг, өөрөөр хэлбэл энэ нь эрчим хүч зарцуулдаг процесс юм.
ДНХ-ийн хуулбарлах механизм
ДНХ молекул нь өөрөө (хоёр дахин нэмэгдэхгүй) давхар мушгиа юм. Репликацийн явцад түүний хоёр нэмэлт гинжин хэлхээний хоорондох устөрөгчийн холбоо тасардаг. Одоо загвар матрицын үүрэг гүйцэтгэдэг бие даасан гинж бүр дээр нэмэлт нэмэлт гинж бий болсон. Ийнхүү хоёр ДНХ молекул үүсдэг. Тэр тус бүрдээ эхийн ДНХ-ээс нэг гинж авдаг, хоёр дахь нь шинээр нийлэгждэг. Тиймээс ДНХ-ийн репликацийн механизм нь хагас консерватив(нэг гинж хуучин, нэг шинэ). Энэхүү хуулбарлах механизм нь 1958 онд батлагдсан.
ДНХ молекул дахь хэлхээ нь эсрэг параллель байдаг. Энэ нь нэг хэлхээ нь 5" төгсгөлөөс 3" хүртэлх чиглэлд, нэмэлт нь эсрэгээр явдаг гэсэн үг юм. 5 ба 3 тоо нь нуклеотид бүрийн нэг хэсэг болох дезоксирибоз дахь нүүрстөрөгчийн атомын тоог илэрхийлдэг. Нуклеотидууд нь эдгээр атомуудаар фосфодиэфирийн холбоогоор холбогддог. Нэг гинж нь 3 "холболттой, нөгөө нь 5" байдаг тул энэ нь доошоо доошоо байрладаг, өөрөөр хэлбэл нөгөө чиглэлд явдаг. Тодорхой болгохын тулд та ширээн дээр сууж буй нэгдүгээр ангийн сурагч шиг гараа гар дээрээ тавьсан гэж төсөөлж болно.
ДНХ-ийн шинэ хэлхээний өргөтгөлийг гүйцэтгэдэг гол фермент нь зөвхөн нэг чиглэлд үүнийг хийх чадвартай. Тухайлбал: шинэ нуклеотидыг зөвхөн 3" төгсгөлд хавсаргана. Тиймээс синтез нь зөвхөн 5"-аас 3" хүртэлх чиглэлд явж болно.
Гинж нь антипараллель байдаг бөгөөд энэ нь синтез нь өөр өөр чиглэлд явах ёстой гэсэн үг юм. Хэрэв ДНХ-ийн гинж нь эхлээд бүрэн хуваагдаж, дараа нь шинэ нэмэлт гинжийг бий болгосон бол энэ нь асуудал биш байх болно. Бодит байдал дээр гинж нь тодорхой хэмжээгээр ялгаатай байдаг хуулбарлах эхлэлийн цэгүүд, мөн эдгээр газруудад матрицын синтез нэн даруй эхэлдэг.
гэж нэрлэгддэг хуулбарлах сэрээ. Үүний зэрэгцээ нэг эх гинжин хэлхээнд синтез нь салаа салах чиглэлд явагддаг бөгөөд энэ синтез нь тасралтгүй, тасралтгүй явагддаг. Хоёрдахь загвар дээр синтез нь анхны ДНХ-ийн гинжин хэлхээний ялгарах чиглэлээс эсрэг чиглэлд явагддаг. Тиймээс ийм урвуу синтез нь зөвхөн хэсэг хэсгээр нь үргэлжлэх боломжтой бөгөөд үүнийг нэрлэдэг Оказакигийн хэсгүүд. Дараа нь ийм хэлтэрхийнүүд хоорондоо "оёдог".
Тасралтгүй давтагддаг хүүхдийн гинжийг нэрлэдэг тэргүүлэх эсвэл тэргүүлэх. Оказаки хэлтэрхийнүүдээр нийлэгжсэн нь - хоцрогдсон, эсвэл хоцрогдсонУчир нь хуваагдмал хуулбар нь удаан байдаг.
Диаграммд эцэг эхийн ДНХ-ийн хэлхээ нь тэргүүлэх охин хэлхээ нийлэгжсэн чиглэлд аажмаар зөрж байна. Хоцрогдсон гинжин хэлхээний синтез нь салалтын эсрэг чиглэлд явагддаг тул үүнийг хэсэг хэсгээр нь хийх хэрэгтэй.
ДНХ-ийн синтезийн гол ферментийн (полимераз) өөр нэг онцлог нь нийлэгжилтийг өөрөө эхлүүлж чаддаггүй, зөвхөн үргэлжлүүлдэг явдал юм. Түүнд хэрэгтэй праймер эсвэл праймер. Тиймээс жижиг нэмэлт РНХ-ийн бүсийг эхлээд эцэг эхийн хэлхээнд нэгтгэж, дараа нь полимераз ашиглан гинжийг сунгана. Дараа нь праймеруудыг зайлуулж, нүхийг босгоно.
Диаграммд үрийг зөвхөн хоцрогдсон гинж дээр харуулав. Үнэндээ тэд тэргүүлж байна. Гэсэн хэдий ч энд нэг сэрээ тутамд зөвхөн нэг праймер хэрэгтэй.
Эхийн ДНХ-ийн гинж нь төгсгөлөөс нь үргэлж салдаггүй, гэхдээ эхлүүлэх цэг дээр үнэндээ нүд, бөмбөлөг шиг тийм их сэрээ үүсдэггүй.
Бөмбөлөг бүр нь хоёр салаатай байж болно, өөрөөр хэлбэл гинж нь хоёр чиглэлд хуваагдана. Гэсэн хэдий ч тэд зөвхөн нэгийг л хийж чадна. Гэсэн хэдий ч зөрүү нь хоёр чиглэлтэй байвал ДНХ-ийн нэг хэлхээ дээр эхлүүлэх үеэс эхлэн синтез нь урагш ба хойшоо гэсэн хоёр чиглэлд явагдана. Энэ тохиолдолд тасралтгүй синтезийг нэг чиглэлд, Оказаки фрагментийг нөгөө чиглэлд хийнэ.
Прокариот ДНХ нь шугаман биш, харин дугуй бүтэцтэй, зөвхөн нэг хуулбарлах гаралтай байдаг.
Диаграммд ДНХ-ийн эх молекулын хоёр хэлхээг улаан, цэнхэр өнгөөр үзүүлэв. Синтез хийж буй шинэ утаснуудыг тасархай шугамаар харуулав.
Прокариотуудад ДНХ-ийг өөрөө хуулбарлах нь эукариотуудаас хурдан байдаг. Хэрэв эукариотуудын репликацийн хурд секундэд хэдэн зуун нуклеотид байдаг бол прокариотуудад мянга ба түүнээс дээш байдаг.
Хуулбарлах ферментүүд
ДНХ-ийн репликацийг ферментийн цогцолбор гэж нэрлэдэг хариу үйлдэл. Нийт 15 гаруй фермент, репликацийн уураг байдаг бөгөөд хамгийн чухал нь доор жагсаав.
Хуулбарлах гол фермент нь аль хэдийн дурдсан байдаг ДНХ полимераз(үнэндээ хэд хэдэн өөр байдаг), энэ нь гинжин хэлхээний өсөлтийг шууд хэрэгжүүлдэг. Энэ нь ферментийн цорын ганц үүрэг биш юм. Полимераз нь аль нуклеотид төгсгөлд нь нэгдэхийг оролдож байгааг "шалгах" чадвартай. Хэрэв тохиромжгүй бол тэр үүнийг устгана. Өөрөөр хэлбэл, ДНХ-ийн хэсэгчилсэн засвар, өөрөөр хэлбэл хуулбарлах алдааг засах нь синтезийн үе шатанд аль хэдийн явагддаг.
Нуклеоплазмд (эсвэл бактери дахь цитоплазм) олдсон нуклеотидууд нь трифосфат хэлбэрээр байдаг, өөрөөр хэлбэл тэдгээр нь нуклеотид биш, харин дезоксинуклеозид трифосфатууд (dATP, dTTP, dGTP, dCTP) юм. Эдгээр нь ATP-тэй төстэй бөгөөд гурван фосфатын үлдэгдэлтэй бөгөөд хоёр нь макроэргик бондоор холбогддог. Эдгээр холбоо тасрахад маш их энерги ялгардаг. Мөн дезоксинуклеозид трифосфатуудад хоёр холбоо нь макроэргик шинж чанартай байдаг. Полимераз нь сүүлийн хоёр фосфатыг салгаж, ялгарсан энергийг ДНХ полимержих урвалд зарцуулдаг.
Фермент геликазДНХ-ийн матрицын хэлхээг тэдгээрийн хоорондох устөрөгчийн холбоог таслах замаар тусгаарладаг.
ДНХ молекул нь давхар мушгиа тул холбоо тасрах нь улам бүр мушгих болно. Хоёр олсыг бие биенээсээ эргүүлж, нэг талдаа нэг үзүүрийг баруун тийш, нөгөөг нь зүүн тийш татна гэж төсөөлөөд үз дээ. Нэхмэл хэсэг нь улам эрчилсэн болж, илүү нягт болно.
Ийм хурцадмал байдлыг арилгахын тулд салаагүй байгаа давхар мушгиа нь тэнхлэгээ тойрон хурдан эргэлдэж, үүссэн супер ороомгийг "дахин тохируулах" шаардлагатай. Гэсэн хэдий ч энэ нь хэт их эрчим хүч шаарддаг. Тиймээс эсэд өөр механизм хэрэгждэг. Фермент топоизомеразаутаснуудын аль нэгийг тасалж, хоёр дахь нь завсараар дамжуулж, эхнийх нь дахин оёдог. Энэ нь шинээр гарч ирж буй супер ороомогуудыг арилгадаг.
Геликазын үйл ажиллагааны үр дүнд тархсан матрицын ДНХ-ийн хэлхээ нь устөрөгчийн бондтой дахин холбогдохыг оролддог. Үүнээс урьдчилан сэргийлэхийн тулд ДНХ холбогч уураг. Тэд урвалыг хурдасгадаггүй гэдэг утгаараа фермент биш юм. Ийм уургууд нь ДНХ-ийн хэлхээнд бүхэл бүтэн уртаараа наалддаг бөгөөд загвар ДНХ-ийн нэмэлт хэлхээг хаахыг зөвшөөрдөггүй.
Праймеруудыг нэгтгэдэг РНХ примаза. Мөн хасагдсан экзонуклеаз. Праймерыг арилгасны дараа өөр төрлийн полимераз нь "нүх" үүсгэдэг. Гэсэн хэдий ч энэ тохиолдолд ДНХ-ийн бие даасан хэсгүүдийг хооронд нь оёдоггүй.
Синтезжүүлсэн гинжин хэлхээний салангид хэсгүүд нь ийм репликацийн ферментээр хоорондоо холбогддог ДНХ лигаз.