М'язові тканинипоєднує здатність до скорочення.
Особливості будови: скорочувальний апарат, що займає значну частину в цитоплазмі структурних елементів м'язової тканини і складається з актинових та міозинових філаментів, які формують органели спеціального призначення. міофібрили .
Класифікація м'язових тканин
1. Морфофункціональна класифікація:
1) Поперечносмугаста, або смугаста м'язова тканина: скелетна та серцева;
2) Невичерпана м'язова тканина: гладка.
2. Гістогенетична класифікація (залежно від джерел розвитку):
1) Соматичного типу(З міотомів сомітів) - скелетна м'язова тканина (поперечносмугаста);
2) Ціломічного типу(З міоепікардіальної пластинки вісцерального листка спланхнотома) - серцева м'язова тканина (поперечносмугаста);
3) Мезенхімного типу(Розвивається з мезенхіми) - гладка м'язова тканина;
4) Зі шкірної ектодермиі прехордальної платівки- Міоепітеліальні клітини залоз (гладкі міоцити);
5) Нейральногопоходження (з нервової трубки) – міонейральні клітини (гладкі м'язи, що звужують та розширюють зіницю).
Функції м'язової тканини: переміщення тіла або його частин у просторі.
Скелетна м'язова тканина
Смугаста (поперечно-смугаста) м'язова тканинастановить до 40% маси дорослої людини, входить до складу скелетних м'язів, м'язів язика, гортані та ін. Належать до довільних м'язів, оскільки їх скорочення підкоряються волі людини. Саме ці м'язи задіяні під час зайняття спортом.
Гістогенез.Скелетна м'язова тканина розвивається із клітин міотомів міобластів. Розрізняють головні, шийні, грудні, поперекові, крижові міотоми. Вони розростаються у дорзальному та вентральному напрямках. У них рано вростають гілки спинномозкових нервів. Частина міобластів диференціюється дома (утворюють аутохтонную мускулатуру), інші з 3 тижня внутрішньоутробного розвитку мігрують у мезенхіму і, зливаючись друг з одним, утворюють м'язові трубки (міотуби) з великими центрально орієнтованими ядрами. У міотубах відбувається диференціювання спеціальних органел міофібрил. Спочатку вони розташовуються під плазмолемою, а потім заповнюють більшу частину міотуби. Ядра зміщуються до периферії. Клітинні центри та мікротрубочки зникають, грЕПС значно редукується. Така багатоядерна структура називається симпласт , а для м'язової тканини – міосимпласт . Частина міобластів диференціюється в міосателітоцити, які розташовуються на поверхні міосимпластів і згодом беруть участь у регенерації м'язової тканини.
Будова кістякової м'язової тканини
Розглянемо будову м'язової тканини на кількох рівнях організації живого: на органному рівні (м'яз як орган), на тканинному (безпосередньо м'язова тканина), на клітинному (будова м'язового волокна), на субклітинному (будова міофібрили) і на молекулярному рівні (будова актинових та міозинових).
 На картинці:
На картинці:
1 - м'яз литковий (органний рівень), 2 - поперечний зріз м'яза (тканинний рівень) - м'язові волокна, між якими РВСТ: 3 - ендомізій, 4 - нервове волокно, 5 - кровоносна судина; 6 – поперечний зріз м'язового волокна (клітинний рівень): 7 – ядра м'язового волокна – симпласту, 8 – мітохондрія між міофібрилами, синім кольором – саркоплазматичний ретикулум; 9 – поперечний зріз міофібрили (субклітинний рівень): 10 – тонкі актинові нитки, 11 – товсті міозинові нитки, 12 – головки товстих міозинових ниток.
1) Органний рівень: будова м'язи як орган.
Скелетний м'яз складається з пучків м'язових волокон, пов'язаних воєдино системою сполучнотканинних компонентів. Ендомізій– прошарки РВСТ між м'язовими волокнами, де проходять кровоносні судини, нервові закінчення . Перімізій– оточує 10-100 пучків м'язових волокон. Епімізій- Зовнішня оболонка м'язи, представлена щільною волокнистою тканиною.
2) Тканинний рівень: будова м'язової тканини.
 Структурно-функціональною одиницею скелетної поперечно-смугастої м'язової тканини є м'язове волокно- циліндричної форми утворення діаметром 50 мкм і довжиною від 1 до 10-20 см. М'язове волокно складається з 1) міосимпласту(освіта його дивись вище, будова – нижче); 2) дрібних камбіальних клітин – міосателітоцитів, прилеглих до поверхні миосимпласта і які у поглибленнях його плазмолемы, 3) базальної мембрани, якої покрита плазмолема. Комплекс плазмолеми та базальної мембрани називається сарколемма. Для м'язового волокна характерна поперечна смугастість, ядра зміщені на периферію. Між м'язовими волокнами – прошарки РВСТ (ендомізії).
Структурно-функціональною одиницею скелетної поперечно-смугастої м'язової тканини є м'язове волокно- циліндричної форми утворення діаметром 50 мкм і довжиною від 1 до 10-20 см. М'язове волокно складається з 1) міосимпласту(освіта його дивись вище, будова – нижче); 2) дрібних камбіальних клітин – міосателітоцитів, прилеглих до поверхні миосимпласта і які у поглибленнях його плазмолемы, 3) базальної мембрани, якої покрита плазмолема. Комплекс плазмолеми та базальної мембрани називається сарколемма. Для м'язового волокна характерна поперечна смугастість, ядра зміщені на периферію. Між м'язовими волокнами – прошарки РВСТ (ендомізії).
3) Клітинний рівень: будова м'язового волокна (міосимпласту).
Термін «м'язове волокно» має на увазі «міосимпласт», оскільки міосимпласт забезпечує функцію скорочення, міосателітоцити беруть участь тільки в регенерації.
Міосімпласт, Як і клітина, складається з 3-х компонентів: ядра (точніше безліч ядер), цитоплазми (саркоплазма) і плазмолеми (яка покрита базальною мембраною і називається сарколема). Майже весь обсяг цитоплазми заповнений міофібрилами – органелами спеціального призначення, органели загального призначення: грЕПС, аЕПС, мітохондрії, комплекс Гольджі, лізосоми, а також ядра зміщені на периферію волокна.
У м'язовому волокні (міосимпласті) розрізняють функціональні апарати: мембранний, фібрилярний(скоротливий) та трофічний.
Трофічний апаратвключає ядра, саркоплазму та цитоплазматичні органели: мітохондрії (синтез енергії), грЕПС та комплекс Гольджі (синтез білків – структурних компонентів міофібрил), лізосоми (фагоцитоз зношених структурних компонентів волокна).
Мембранний апарат: кожне м'язове волокно покрите сарколеммою, де розрізняють зовнішню базальну мембрану та плазмолему (під базальною мембраною), яка утворює вп'ячування ( Т-трубочки). До кожної Т-Трубочці примикають по дві цистерни тріаду: дві L-трубочки (цистерни аЕПС) та одна Т-трубочка (вп'ячування плазмолеми). У цистернах аЕПС концентруються Са 2+, необхідний при скороченні. До плазмолеми зовні належать міосателітоцити. При пошкодженні базальної мембрани запускається мітотичний цикл міосателітоцитів.
Фібрилярний апарат. Більшу частину цитоплазми смугастих волокон займають органели спеціального призначення – міофібрили, орієнтовані поздовжньо, що забезпечують скорочувальну функцію тканини.
4) Субклітинний рівень: будова міофібрили.
При дослідженні м'язових волокон і міофібрил під світловим мікроскопом відзначається чергування в них темних і світлих ділянок – дисків. Темні диски відрізняються подвійним променезаломленням і називаються анізотропними дисками, або А- Дисками. Світлі диски не мають подвійного променезаломлення і називаються ізотропними, або I-Дисками.
У середині диска Ає більш світла ділянка – Н-Зона, де містяться тільки товсті нитки білка міозину. У середині Н-Зони (означає і А-диска) виділяється темніша М-Лінія, що складається з міомезину (необхідний для складання товстих ниток та їх фіксації при скороченні). У середині диска Iрозташована щільна лінія Zяка побудована з білкових фібрилярних молекул. Z-лінія з'єднана з сусідніми міофібрилами за допомогою білка десміну, і тому всі названі лінії та диски сусідніх міофібрил збігаються і створюється картина поперечносмугастої смугастість м'язового волокна.
Структурною одиницею міофібрили є саркомір (S) — це пучок міофіламентів укладений між двома Z-лініями. Міофібрилла складається з безлічі саркомірів. Формула, що описує структуру саркомера:
S = Z 1 + 1/2 I 1 + А + 1/2 I 2 + Z 2

5) Молекулярний рівень: будова актинових і міозинових філаментів .
Під електронним мікроскопом міофібрили представляють агрегати з товстих, або міозинових, і тонких, або актинових, Філаменти. Між товстими філаментами розташовуються тонкі філаменти (діаметр 7-8 нм).
Товсті філаменти, або міозинові нитки,(Діаметр 14 нм, довжина 1500 нм, відстань між ними 20-30 нм) складаються з молекул білка міозину, що є найважливішим скоротливим білком м'язи, по 300-400 молекул міозину в кожній нитці. Молекула міозину – це гексамер, що складається з двох важких та чотирьох легких кіл. Важкі ланцюги є дві спірально закручені поліпептидні нитки. Вони несуть на своїх кінцях кулясті голівки. Між головкою та важким ланцюгом знаходиться шарнірна ділянка, за допомогою якої головка може змінювати свою конфігурацію. В області головок – легкі ланцюги (по два на кожній). Молекули міозину укладені в товстій нитці таким чином, що їхні головки звернені назовні, виступаючи над поверхнею товстої нитки, а важкі ланцюги утворюють стрижень товстої нитки.
Міозин має АТФ-азну активність: енергія, що вивільняється, використовується для м'язового скорочення.
Тонкі філаменти, або актинові нитки,(діаметр 7-8 нм), утворені трьома білками: актином, тропоніном та тропоміозином. Основним за масою білком є актин, який утворює спіраль. Молекули тропоміозину розташовуються у жолобку цієї спіралі, молекули тропоніну розташовуються вздовж спіралі.
Товсті нитки займають центральну частину саркомера. А-диск, тонкі займають I- диски та частково входять між товстими міофіламентами. Н-Зона складається тільки з товстих ниток.
В спокої взаємодія тонких та товстих ниток (міофіламентів)неможливо, т.к. міозин-зв'язувальні ділянки актину заблоковані тропоніном та тропоміозином. При високій концентрації іонів кальцію конформаційні зміни тропоміозину призводять до розблокування міозин-зв'язувальних ділянок молекул актину.
Двигуна іннервація м'язового волокна. Кожне м'язове волокно має власний апарат іннервації (моторна бляшка) і оточене мережею гемокапілярів, що знаходяться в прилеглій РВСТ. Цей комплекс називається міон.Група м'язових волокон, що іннервуються одним мотонейроном, називається нервово-м'язової одиницею.М'язові волокна у разі можуть розташовуватися не поруч (одне нервове закінчення може контролювати від однієї до десятків м'язових волокон).
При надходженні нервових імпульсів за аксонами рухових нейронів відбувається скорочення м'язового волокна.
Скорочення м'яза
При скороченні м'язові волокна коротшають, але довжина актинових та міозинових філаментів у міофібрилах не змінюється, а відбувається їх рух один щодо одного: міозинові нитки всуваються у простори між актиновими а, актинові – між міозиновими. Внаслідок цього зменшується ширина I-диска, H-смужки та зменшується довжина саркомера; ширина А-Диска не змінюється.
Формула саркомера при повному скороченні: S = Z 1 + А+ Z 2
Молекулярний механізм м'язового скорочення
1. Проходження нервового імпульсу через нервово-м'язовий синапс та деполяризація плазмолеми м'язового волокна;
2. Хвиля деполяризації проходить по Т-трубочкам (вп'ячування плазмолеми) до L-трубочок (цистерни саркоплазматичного ретикулуму);
3. Відкриття кальцієвих каналів у саркоплазматичному ретикулумі та вихід іонів Са 2+ у саркоплазму;
4. Кальцій дифундує до тонких ниток саркомера, зв'язується з тропоніном С, призводячи до конформаційних змін тропоміозину та звільняючи активні центри для зв'язування міозину та актину;
5. Взаємодія міозинових головок з активними центрами на молекулі актину з утворенням актино-міозинових «містків»;
6. Міозинові головки «крочать» актином, утворюючи в ході переміщення нові зв'язки актину і міозину, при цьому актинові нитки підтягуються в простір між міозиновими нитками до M-лінії, зближуючи дві Z-Лінії;
7. Розслаблення: Са 2+ -АТФ-аза саркоплазматичного ретикулуму закачує Са 2+ із саркоплазми до цистерн. У саркоплазмі концентрація Са 2+ стає низькою. Розриваються зв'язки тропоніну Зз кальцієм, тропоміозин закриває міозин-зв'язуючі ділянки тонких ниток і перешкоджає їх взаємодії з міозином.
Кожен рух головки міозину (приєднання до актину та від'єднання) супроводжується витратою енергії АТФ.
Чутлива іннервація(нервово-м'язові веретени). Інтрафузальні м'язові волокна разом із чутливими нервовими закінченнями формують нервово-м'язові веретени, які є рецепторами скелетного м'яза. Зовні сформовано капсулу веретену. При скороченні поперечно-смугастих (смугастих) м'язових волокон змінюється натяг сполучно-тканинної капсули веретена і відповідно змінюється тонус інтрафузальних (розташованих під капсулою) м'язових волокон. Формується нервовий імпульс. При надмірному розтягуванні м'яза виникає відчуття болю.
Класифікація та типи м'язових волокон
1. За характером скорочення: фазні та тонічнім'язові волокна. Фазні здатні здійснювати швидкі скорочення, але не можуть довго утримувати досягнутий рівень укорочення. Тонічні м'язові волокна (повільні) забезпечують підтримку статичної напруги або тонусу, що відіграє роль у збереженні певного положення тіла у просторі.
2. За біохімічними особливостями та кольором виділяють червоні та білі м'язові волокна. Колір м'яза обумовлений ступенем васкуляризації та вмістом міоглобіну. Характерною особливістю червоних м'язових волокон є наявність численних мітохондрій, ланцюги яких розташовуються між міофібрилами. У білих м'язових волокнах мітохондрій менше і вони знаходяться рівномірно в саркоплазмі м'язового волокна.
3. За типом окисного обміну : оксидативні, гліколітичні та проміжні. Ідентифікація м'язових волокон заснована на виявленні активності ферменту сукцинатдегідрогенази (СДГ), яка є маркером для мітохондрій та циклу Кребса. Активність цього ферменту свідчить про напруженість енергетичного метаболізму. Виділяють м'язові волокна А-типу (гліколітичні) з низькою активністю СДГ, З-Тип (оксидативні) з високою активністю СДГ. М'язові волокна У-Типу займають проміжне положення. Перехід м'язових волокон від А-Типу в З-Тип маркує зміни від анаеробного гліколізу до метаболізму, що залежить від кисню.
У спринтерів (спортсменів, коли потрібне швидке недовге скорочення, культуристів) тренування та харчування спрямоване на розвиток гліколітичних, швидких, білих м'язових вовкон: у них багато запасів глікогену та енергія видобувається переважно анаеолбним шляхом (біле м'ясо у курки). У стаєрів (спортсменів - марафонців, у тих видах спорту, де необхідна витривалість) переважають оксидативні, повільні, червоні волокна в м'язах - у них багато мітохондрій для аеробного гліколізу, кровоносних судин (потрібний кисень).
4. У смугастих м'язах розрізняють два види м'язових волокон: екстрафузальні, які переважають і обумовлюють власне скорочувальну функцію м'язу та інтрафузальні, що входять до складу пропріоцепторів - нервово-м'язових веретен.
Факторами, що визначають структуру та функцію скелетного м'яза є вплив нервової тканини, гормональний вплив, місце розташування м'яза, рівень васкуляризації та рухової активності.
Серцева м'язова тканина
Серцева м'язова тканина знаходиться в м'язовій оболонці серця (міокард) і в гирлах пов'язаних з ним великих судин. Має клітинний тип будови та основною функціональною властивістю служить здатність до спонтанних ритмічних скорочень (мимовільні скорочення).
Розвивається з міоепікардіальної платівки (вісцеральний листок спланхнотома мезодерми в шийному відділі), клітини якої розмножуються мітозом, а потім диференціюються. У клітинах з'являються міофіламенти, які формують міофібрили.
Будова. Структурна одиниця серцевої м'язової тканини – клітина кардіоміоцит.Між клітинами знаходяться прошарки РВСТ із кровоносними судинами та нервами.
Типи кардіоміоцитів : 1) типові (робітники, скорочувальні), 2) атипові(провідні), 3) секреторні.
Типові кардіоміоцити
Типові (робочі, скорочувальні) кардіоміоцити- Клітини циліндричної форми, довжиною до 100-150 мкм і діаметром 10-20 мкм. Кардіоміоцити утворюють основну частину міокарда, з'єднані один з одним у ланцюжка основами циліндрів. Ці зони називають вставними дисками, в яких виділяють десмосомальні контакти та нексуси (щілинні контакти). Десмосоми забезпечують механічне зчеплення, яке перешкоджає розбіжності кардіоміоцитів. Щелевидные контакти сприяють передачі скорочення від одного кардіоміоциту до іншого.
Кожен кардіоміоцит містить одне або два ядра, саркоплазму та плазмолему, оточену базальною мембраною. Розрізняють функціональні апарати, такі ж, як у м'язовому волокні: мембранний, фібрилярний(скоротливий), трофічний,а також енергетичний.
Трофічний апарат включає ядро, саркоплазму та цитоплазматичні органели: грЕПС та комплекс Гольджі (синтез білків – структурних компонентів міофібрил), лізосоми (фагоцитоз структурних компонентів клітини). Кардіоміоцити, як і олокна скелетної м'язової тканини, характеризуються наявністю в їх саркоплазмі залізовмісного кисень-зв'язуючого пігменту міоглобіну, що надає їм червоного кольору і схожого за будовою та функцією з гемоглобіном еритроцитів.
Енергетичний апарат представлений мітохондріями та включеннями, розщеплення яких забезпечує отримання енергії. Мітохондрії численні, лежать рядами між фібрилами, біля полюсів ядра та під сарколемою. Енергія, необхідна кардіоміоцитам, виходить шляхом розщеплення: 1) основного енергетичного субстрату цих клітин жирних кислот, які депонуються у вигляді тригліцеридів у ліпідних краплях; 2) глікогену, що знаходиться в гранулах, розташованих між фібрилами.
Мембранний апарат : кожна клітина покрита оболонкою, що складається з комплексу плазмолеми та базальної мембрани. Оболонка утворює вп'ячування ( Т-трубочки). До кожної Т-трубочці примикає одна цистерна (на відміну м'язового волокна – там 2 цистерни) саркоплазматичного ретикулуму(видозмінена аЕПС), утворюючи діаду: одна L-трубочка (цистерна аЕПС) та одна Т-трубочка (вп'ячування плазмолеми). У цистернах аЕПС іони Са 2+ накопичуються не так активно, як у м'язових волокнах.
Фібрилярний (скорочувальний) апарат . Більшу частину цитоплазми кардіоміоциту займають органели спеціального призначення – міофібрили, орієнтовані поздовжньо і розташовані по периферії клітини. При розслабленні іони кальцію виділяються в саркоплазму з низькою швидкістю, що забезпечує автоматизм і часті скорочення кардіоміоцитів. Т-трубочки широкі та утворюють діади (одна Т-трубочка та одна цистерна мережі), які сходяться в області Z-Лінії.
Кардіоміоцити, зв'язуючись за допомогою вставних дисків, утворюють скорочувальні комплекси, які сприяють синхронізації скорочення, між кардіоміоцитами сусідніх скорочувальних комплексів утворюються бічні анастомози.
Функція типових кардіоміоцитів: забезпечення сили скорочення серцевого м'яза
Провідні (атипові) кардіоміоцитиволодіють здатністю до генерації та швидкого проведення електричних імпульсів. Вони утворюють вузли та пучки провідної системи серця і поділяються на кілька підтипів: пейсмекери (в синоатріальному вузлі), перехідні (в атріо-вентрикулярному вузлі) та клітини пучка Гіса та волокон Пуркіньє. Провідні кардіоміоцити характеризуються слабким розвитком скорочувального апарату, світлою цитоплазмою та великими ядрами. У клітинах немає Т-трубочок і поперечної смугастість, оскільки міофібрили розташовані невпорядковано.
Функція атипових кардіоміоцитів- генерація імпульсів та передача на робочі кардіоміоцити, забезпечуючи автоматизм скорочення міокарда.
Секреторні кардіоміоцити
Секреторні кардіоміоцити перебувають у передсердях, переважно у правому; характеризуються відростчастою формою та слабким розвитком скорочувального апарату. У цитоплзмі, поблизу полюсів ядра – секреторні гранули, що містять натріуретичний фактор, або атріопептин(Гормон, що регулює артеріальний тиск). Гормон викликає втрату натрію та води із сечею, розширення судин, зниження тиску, пригнічення секреції альдостерону, кортизолу, вазопресину.
Функція секреторних кардіоміоцитів: ендокринна.
Регенерація кардіоміоцитів.Для кардіоміоцитів характерна лише внутрішньоклітинна регенерація. Кардіоміоцити не здатні до поділу, у них відсутні камбіальні клітини.
Гладка м'язова тканина
Гладка м'язова тканина утворює стінки внутрішніх порожнистих органів, судин; характеризується відсутністю смугастість, мимовільними скороченнями. Іннервація здійснюється вегетативною нервовою системою.
Структурно-функціональна одиниця невикресленої гладкої м'язової тканини гладенька м'язова клітина (ГМК), або гладкий міоцит.Клітини мають веретеноподібну форму довжиною 20-1000 мкм та товщиною від 2 до 20 мкм. У матці клітини мають витягнуту відростчасту форму.
Гладкий міоцит
Гладкий міоцит складається з розташованого в центрі ядра паличкоподібної форми, цитоплазми з органелами та сарколеми (комплекс плазмолеми та базальної мембрани). У цитоплазмі біля полюсів знаходиться комплекс Гольджі, багато мітохондрій, рибосом, розвинений саркоплазматичний ретикулум. Міофіламенти розташовані косо або вздовж поздовжньої осі. У ГМК актинові та міозинові філаменти не формують міофібрил. Актинових ниток більше і вони прикріплюються до щільних тільців, які утворені спеціальними білками, що зшивають. Поруч із актиновими нитками розташовуються мономери міозину (мікроміозин). Маючи різну довжину, вони значно коротші за тонкі нитки.
Скорочення гладких м'язових клітинздійснюється при взаємодії актинових філаментів та міозину. Сигнал, що йде нервовими волокнами, зумовлює виділення медіатора, що змінює стан плазмолеми. Вона утворює колбоподібні вп'ячування (кавеоли), де концентруються іони кальцію. Скорочення ГМК індукується припливом іонів кальцію в цитооплазму: кавеоли відшнуровуються разом із іонами кальцію потрапляють у клітину. Це призводить до полімеризації міозину та взаємодії його з актином. Актинові нитки та щільні тільця зближуються, зусилля передається на сарколему та ГМК коротшає. Міозин у гладких міоцитах здатний взаємодіяти з актином лише після фосфорилювання його легких ланцюгів спеціальним ферментом – кіназою легких ланцюгів. Після припинення сигналу іони кальцію залишають кавеоли; міозин деполяризується, втрачає спорідненість до актину. В результаті комплекси міофіламентів розпадаються; скорочення припиняється.
Особливі типи м'язових клітин
Міоепітеліальні клітини є похідними ектодерми, що не мають смугастість. Оточують секреторні відділи та вивідні протоки залоз (слинних, молочних, слізних). Із залізистими клітинами вони пов'язані десмосомами. Скорочуючись, сприяють виділенню секрету. У кінцевих (секреторних) відділах форма клітин відросткова, зірчаста. Ядро в центрі, в цитоплазмі, переважно у відростках, локалізовані міофіламенти, які утворюють скорочувальний апарат. У цих клітинах є і цитокератинові проміжні філаменти, що підкреслює їхню схожість з епітеліоцитами.
Міонейральні клітини розвиваються з клітин зовнішнього шару очного келиха і утворюють м'яз, що звужує зіницю і м'яз, що розширює зіницю. За будовою перший м'яз подібний до ГМК мезенхімного походження. М'яз, що розширює зіницю утворена відростками клітин, що розташовуються радіально, а ядросодержащая частина клітини знаходиться між пігментним епітелієм і стромою райдужної оболонки.
Міофібробласти відносяться до пухкої сполучної тканини і є видозміненими фібробластами. Вони виявляють властивості фібробластів (синтезують міжклітинну речовину) і гладких міоцитів (мають виражені скорочувальні властивості). Як варіант цих клітин можна розглядати міоїдні клітини у складі стінки звивистого насіннєвого канальця яєчка та зовнішнього шару теки фолікула яєчника. При загоєнні рани частина фібробластів синтезує гладком'язові актини та міозини. Міофібробласти забезпечують стягування країв рани.
Ендокринні гладкі міоцити – це видозмінені ГМК, що становлять основний компонент юкстагломерулярного апарату нирок. Вони знаходяться в стінці артеріол ниркового тільця, мають добре розвинений синтетичний апарат і скорочувальний редукований. Продукують фермент ренін, що знаходиться в гранулах і потрапляє в кров механізмом екзоцитозу.
Регенерація гладкої м'язової тканини.Гладкі міоцити характеризуються внутрішньоклітинною регенерацією. При підвищенні функціонального навантаження відбувається гіпертрофія міоцитів та в деяких органах гіперплазія (клітинна регенерація). Так, при вагітності гладко-м'язові клітини матки можуть збільшуватись у 300 разів.
Поперечнополосатая скелетна м'язова тканина утворює м'язи голови, тулуба, кінцівок, глотки, гортані, жувальних м'язів, язика, краніального відділу стравоходу. Іннервація за рахунок соматичної нервової системи дозволяє тварині здійснювати довільні скорочення скелетних м'язів тіла. Структурно-функціональна одиниця скелетної м'язової тканини – м'язове волокно (міон), що розвивається з міотомів сегментованого відділу (сомітів) мезодерми. Гістогенез м'язових волокон: міобластична, міосимпластична, м'язових трубочок, дефінітивних м'язових волокон (рис. 42).
Міобластична стадіяхарактеризується тим, що на ранній стадії розвитку міотоми складаються з щільно розташованих епітеліоподібних м'язових клітин, що мають великі ядра та тонковолокнисту структуру цитоплазми. Епітеліоподібні м'язові клітини диференціюються в проміобласти, а потім у міобласти, які активно діляться і переміщуються як єдине ціле потоками до ділянок розташування майбутніх м'язів. Одна частина міобластів не бере участі в утворенні м'язових волокон і диференціюється в сателіти - камбіальні клітини, які розташовуються між базальною мембраною та сарколемою. Визначальним моментом переходу до міосимпластичної стадіїє званий диференціальний мітоз, тобто. блокується репродукція міобластів, які набувають здатності до злиття в симпласти, за рахунок розподілу ядер без поділу цитоплазми. Тому спостерігається утворення багатоядерних структур, у загальній масі цитоплазми є поодинокі скорочувальні нитки - міофібрили, яких
Мал. 42
а- міобластична стадія; 6 - Міосимпластична стадія; в- стадія м'язових трубочок; г- стадія формування дефінітивних м'язових волокон;
7 - епітеліоподібні клітини міотома; 2 - проміобласти; 3 - міобласти; 4 - Мітоз; 5 - злиття міобластів та початок синтезу міофібрил; б - утворення міосимпласту
стає дедалі більше. Стадія м'язових трубочокхарактеризується тим, що багатоядерні структури набувають подовжену трубчасту форму, в центральній частині яких ряд розміщуються численні ядра, на периферії диференціюються міофібрили. Після того, як більшість м'язових трубочок заповнюється мі- офібрилами, встановлюються нейром'язові контакти. Формування дефінітивних м'язових волоконсупроводжується різким збільшенням числа міофібрил, що займають центральне положення, тоді як численні ядра переміщаються на периферію та розташовуються під плазмолемою.
У постнатальний період зростання м'язів зумовлений збільшенням числа, а потовщенням волокон, тобто. міофіламенты будуються на поверхні вже наявних міофібрил. М'язові волокна ростуть у довжину за рахунок прибудови нових саркомерів, при пошкодженні регенерують за рахунок камбіальних клітин - сателітів, здатних диференціюватися в міобласти і далі, як це відбувається в ембріогенезі.
Структурно-функціональна організація м'язового волокна.М'язове волокно має вигляд довгого тонкого циліндра, довжина якого може досягати 13-15 см, діаметр 10-150 мкм. Зовні волокно оточене оболонкою – сарколеммою (від грец. sarcos – м'ясо; lemma – оболонка), що складається з двох шарів, утворених мембранами. Зовнішня (базальна) та внутрішня (плазматична) мембрани розділені простором 10-25 нм. Базальна мембрана, розташована між пухкою волокнистою сполучною тканиною та м'язовим волокном, відіграє роль посередника і служить місцем прикріплення колагенових та еластичних волокон. Плазматична мембрана (плазмолема)м'язового волокна безпосередньо обмежує вміст саркоплазми, виконує функцію бар'єру, що забезпечує вибірковий обмін речовин між м'язовим волокном та навколишнім середовищем. Цитоплазма м'язового волокна називається саркоплазмою. Між структурами волокна розташована основна речовина саркоплазми. саркоплазматичний матрикс,що складається з глобулярних білків та пігментного білка - міоглобіну, здатного зв'язувати кисень. У саркоплазмі численні ядра розташовуються безпосередньо під сарколеммою, розмір і форма ядер залежить від ступеня скорочення м'язового волокна. Між міофібрилами знаходиться безліч мітохондрій. саркос.Цитоплазма - саркоплазма м'язового волокна містить мережу внутрішніх мембран - саркоплазматичний ретикулум.Поперек волокна, між міофібрилами проходить система трубочок. Т-система,пов'язана із сарколеммою. Комплекс з однієї Г-трубочки, саркоплазматичної мережі та термінальних цистерн називається тріадою.Тріада бере участь у просуванні хвиль деполяризації, акумуляції та вивільнення іонів кальцію. Через війну концентрація цих іонів у саркоплазмі знижується чи підвищується, що, своєю чергою, впливає активність АТФ-азы, отже, на скорочувальну функцію м'язового волокна.
Скоротливий апаратявляє собою поздовжньо орієнтовані білкові скорочувальні нитки. міофібрили,мають вичерчену структуру через наявність світлих і темних дисків, що чергуються. Ці диски по-різному заломлюють поляризоване світло: світлі, ізотропні 1-дискимають одинарний; а темні, анізотропні A-диски- подвійним променезаломленням. У темних дисках знаходяться товсті міозинові нитки, що містять світлу Я-зону, у середині якої проходить М-лінія. Світлі диски містять тонкі актинові нитки, які перетинає Z-лінія посередині. Ділянка міофібрили між Z-лініями. саркомір,отже, кожен саркомір містить один Л-диск і дві половинки I-диска. Саркомір є структурно-функціональною одиницею міофібрили, один кінець тонкої (актинової) нитки прикріплений до
Z-лінії, а інший спрямований на середину саркомера, тобто. тонкі нитки проходять між товстими нитками (рис. 43).

Мал. 43
- 1 – ядро; 2 - міофібрили; 3 - саркоплазма; 4 - саркомір; 5 - сухожилля;
- 6 – оболонка волокна; А- Анізотропний диск; J- Ізотропний диск; Z – темна смужка; Н- Світла смужка
Структура м'язових волокон у складі м'яса та м'ясних виробів.
М'язові волокна у складі м'яса мають гофровану та складчасту форму. Характер складчастості залежить від ступеня скорочення: гофровані волокна мають слабке скорочення або розслаблені, а розташовані поруч із ними прямі волокна сильно скорочені. Ступінь скорочення м'язових волокон визначають за наявністю вузлів скорочення, що являють собою характерну структуру, що виникає в поперечносмугастих м'язових волокнах всіх видів сільськогосподарських тварин. Вузли скорочення характеризуються сильним ступенем скорочення міофібрил; ступінь їх скорочення по довжині вузла часто розподіляється закономірно: найвищий ступінь скорочення - в області, розташованій ближче до центру вузла.
У морфологічному відношенні м'ясо являє собою тканинний комплекс, до складу якого входять м'язова, сполучна (пухка волокниста, жирова, кісткова) тканини, кровоносні та лімфатичні судини, нервові волокна та нервові закінчення. Після забою тварини із припиненням обміну речовин основним біохімічним процесом при переробці сировини є автоліз (від грец. autos – сам, lisis – розчинення), основу якого складають ферментативні процеси. Сукупність змін найважливіших властивостей, зумовлених розвитком автолізу, в результаті яких м'ясо набуває ніжної консистенції та соковитості, добре виражений специфічний запах і смак обумовлює дозрівання м'яса. Парне м'ясо- Початкова структура, з якою можна порівнювати всі наступні зміни в м'ясі. М'язові волокна парного м'яса, як згадувалося, мають гофровану і складчасту форму;
ступінь скорочення м'язових волокон можна визначати за наявності вузлів скорочення, і найвищий ступінь скорочення спостерігається в області, розташованій ближче до центру вузла. Ця маса в центрі вузлів зазвичай стає місцем роз'єднання волокна на окремі фрагменти, кількість яких збільшується, якщо по довжині вузли скорочення досить великі. У охолодженому м'ясіскладчастість м'язових волокон майже повністю зникає, а при заморожуванні м'яса зміни характеризуються появою нового структурного компонента – водних кристалів – та зміною загального виду та товщини м'язових волокон. Кристали в м'ясі утворюються за рахунок перенесення рідини, що кристалізується, з тканинного соку. Енергійні процеси кристалізації є причиною пухкого та хаотичного розташування м'язових волокон, їх фрагментації та деформації.
Препарат «Поперечносмугасті м'язи язика кролика»(Забарвлення залізним гематоксиліном і сумішшю Маллорі). На гістологічних препаратах виявляються поперечносмугасті волокна, обмежені оболонкою. При слабкому збільшенні мікроскопа (х10) виявляються волокна в поздовжньому, поперечному та косому перерізі. Елементи пухкої волокнистої сполучної тканини, розташовані зовні від базальної мембрани, утворюють прошарки - ендомізій, що оточують окремі м'язові волокна, поєднуючи їх у первинні пучки, що сприяє інтеграції зусиль при скороченні. Виявляються також більші прошарки пухкої волокнистої неоформленої сполучної тканини. перимизій,навколишні м'язові пучки різної величини. У складі пухкої волокнистої неоформленої сполучної тканини виявляються колагенові та еластичні волокна, скупчення жирових клітин, кровоносні судини, нервові волокна (рис. 44).

Мал. 44
- 7 - м'язові волокна в поздовжньому та поперечному перерізі; 2,3 - прошарку пухкої волокнистої неоформленої сполучної тканини; 4 - жирові клітини;
- 5 – кровоносні судини
Препарат "Мікроскопічна структура яловичини"(Забарвлення залізним гематоксиліном). На препараті при слабкому збільшенні мікроскопа (х10) виявляються м'язові волокна в поздовжньому та поперечному перерізі. Між м'язовими волокнами проходять дуже тонкі прошарки пухкої волокнистої сполучної тканини – ендомізій. Декілька волокон об'єднані в м'язові пучки різної величини, між цими пучками виявляються прошарки пухкої волокнистої сполучної тканини - перимизій. Ендомізій і перимизій складаються з колагенових волокон різної міцності, що утворюють більш менш складні сплетення, і містять різну кількість еластичних волокон (рис. 45).
Препарат «Мікроструктура м'яса індички»(Забарвлення залізним гематоксиліном). На гістологічних препаратах виявляються поперечносмугасті волокна, обмежені оболонкою. При слабкому збільшенні мікроскопа (х10) виявляються волокна в поздовжньому поперечному перерізі. У поздовжньому перерізі волокна мають вигляд циліндрів, а поперечному перерізі - округлу чи овальну форму. Важливою диференціальною ознакою, що відрізняє м'язи птиці від м'язів ссавців, є розташування ядер у м'язовому волокні: у птахів ядра розташовані в центральній частині, а у ссавців – по периферії волокна (рис. 46).

Мал. 45 . Мікроструктура яловичини: а- Поздовжній зріз м'язових волокон парного м'яса; б, в- поздовжній та поперечний зрізи м'язових волокон замороженого м'яса (забарвлення гематоксиліном;
бл. х7; про. х40)

Мал. 46
бл. х7; про. х40)
Препарат «Поперечносмугасті м'язи коропа»(Забарвлення залізним гематоксиліном). При слабкому збільшенні мікроскопа (х10) виявляються поперечносмугасті волокна, об'єднані в сегменти - міотоми, які розділені сполучнотканинними перегородками - міосептами (рис. 47, а).У поздовжньому перерізі волокна мають вигляд циліндрів, ядра розташовані по всій товщі м'язового волокна. Елементи пухкої волокнистої сполучної тканини оточують як окремі м'язові волокна, і пучки м'язових волокон. У складі сполучної тканини виявляються колагенові та еластичні волокна, скупчення жирових клітин, кровоносні судини, нервові волокна.
Препарат «Поперечносмугасті м'язи жаби»(Забарвлення залізним гематоксиліном). При слабкому збільшенні мікроскопа (х 10) виявляються поперечно смугасті волокна, з центрально розташованими ядрами (рис. 47, б).Відмінна особливість від попередньо-

Мал. 47а- коропа; б- жаби; (забарвлення гематоксиліном та еозином; бл. х7; об. х40) дуючих препаратів: волокна дуже щільно прилягають один до одного, між ними мало елементів пухкої волокнистої сполучної тканини.
Препарат «Поперечносмугаста м'язова тканина при контамінації бактеріями».При контамінації м'яса бактеріями спостерігається так зване бактеріальне псування. В результаті розвитку мікроорганізмів відбувається розпад білка з утворенням продуктів гідролізу, що істотно впливають на органолептичні показники та харчову цінність м'яса. Мікроорганізми, що розвиваються в кислому середовищі, зрушують pHу лужний бік, готують умови для життєдіяльності інших мікроорганізмів. Швидкість псування залежить від температури та вологості навколишнього середовища, стану «кірочки підсихання» та виду бактерій. Як правило, бактеріальне псування починається на поверхні, а потім проникає в товщу м'яса. На гістологічних препаратах дифузно розташовані бактерії виявляються на поверхні м'язових волокон у прошарках пухкої волокнистої сполучної тканини (рис. 48).
У виробах з м'яса заміну м'яса забійних тварин м'ясом інших хребетних тварин виявляють за становищем ядер у м'язовому волокні: у амфібій, риб, птахів ядра розташовані в центральній частині, тоді як у ссавців - по периферії волокна. При використанні м'ясного обрізу, м'яса голів, губ, вух виявляються слинні залози, фрагменти хрящів, багатошаровий плоский епітелій слизової оболонки ротової порожнини. М'ясні консерви можуть містити значну кількість сполучної тканини, добавки рослинних білків і крохмалю, м'язова тканина може замінюватися субпродуктами. Для збільшення

Мал. 48 . Мікроструктура яловичини при контамінації бактеріями: а- Мікроорганізми на поверхні м'язових волокон; б- мікроорганізми у прошарках пухкої волокнистої сполучної тканини обсягів виробництва, раціонального використання сировини, стабілізації якості ковбасних виробів поряд із традиційною сировиною широко використовуються субпродукти, рослинні білки та інші компоненти рослинного та тваринного походження. Наприклад, соєвий білок має вигляд великих фрагментів із щільною волокнистою структурою, що фарбується кислими барвниками. Клітини рослинної тканини великі, чітко окреслені, мають целюлозну оболонку і слабо сприймають гістологічні барвники.
Препарат «Ковбаса варена»(забарвлення гематоксиліном та еозином). При мікроскопії виявляються дрібнозерниста маса аморфної світлішої речовини; окремі фрагменти м'язових волокон; краплі жирової речовини із зруйнованих жирових клітин, при використанні порошкоподібних прянощів виявляються окремі фрагменти або групи клітин. В аморфній речовині зустрічаються досить великі вакуолі (порожнечі овальної та округлої форми) – ділянки, де були повітряні бульбашки, що виникають у процесі технологічної обробки. Технологічні процеси (подрібнення, посол, варіння) відбиваються на ступені деструкції м'язових волокон, розподілі зернистої маси, розмірі та форми вакуолей та розподілі жирової фракції, наявність мікроорганізмів (рис. 49).

Мал. 49а- Контроль: 1 - Оболонка; 2 - Фрагменти м'язових волокон; 3 - Вакуолі;
4 - жирові клітини; б - досвід (контамінація мікроорганізмами) (забарвлення гематоксиліном та еозином; бл. х7; об. х40)
Контрольні питання
- 1. Дайте характеристику поперечносмугастої кістякової м'язової тканини.
- 2. Які основні етапи гістогенезу м'язових волокон?
- 3. Яка структура м'язових волокон?
- 4. Яку структуру мають міофібрили?
- 5. Охарактеризуйте зміни м'язової тканини у процесі технологічної обробки.
- 6. Дайте характеристику мікроструктурі м'яса та м'ясних виробів.
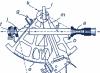
У хребетних тварин та людини розрізняють три різні за будовою групи м'язів:
- поперечно-смугасті м'язи скелета;
- поперечно-смугастий м'яз серця;
- гладкі м'язи внутрішніх органів, судин та шкіри.
Мал. 1. Види м'язів людини
Гладкі м'язи
З двох видів м'язової тканини (поперечно-смугастої та гладкої) гладка м'язова тканина знаходиться на нижчому ступені розвитку і властива нижчим тваринам.
Утворюють м'язовий шар стінок шлунка, кишечника, сечоводів, бронхів, кровоносних судин та інших порожнистих органів. Вони складаються з веретеноподібних м'язових волокон і не мають поперечної смугастість, тому що міофібрили в них розташовані менш впорядковано. У гладких м'язах окремі клітини з'єднуються між собою спеціальними ділянками зовнішніх мембран. нексусами. За рахунок цих контактів потенціали дії поширюються з м'язового волокна на інше. Тому в реакцію збудження швидко залучається весь м'яз.
Гладкі м'язи здійснюють рух внутрішніх органів, кровоносних та лімфатичних судин. У стінках внутрішніх органів вони, як правило, розташовуються у вигляді двох шарів: внутрішнього кільцевого та зовнішнього поздовжнього. У стінках артерії вони утворюють спіралеподібні структури.
Характерною особливістю гладких м'язів є їхня здатність до спонтанної автоматичної діяльності (м'язи шлунка, кишечника, жовчного міхура, сечоводів). Ця властивість регулюється нервовими закінченнями. Гладкі м'язи пластичні, тобто. здатні зберігати надану розтягуванням довжину без зміни напруги. Скелетний м'яз, навпаки, має малу пластичність і цю різницю легко встановити в наступному досвіді: якщо розтягнути за допомогою вантажів і гладкий і поперечно-смугастий м'язи і зняти вантаж, то скелетний м'яз відразу ж після цього коротшає до початкової довжини, а гладкий м'яз довгий час може перебувати в розтягнутому стані.
Така властивість гладких м'язів має велике значення для функціонування внутрішніх органів. Саме пластичність гладких м'язів забезпечує лише невелику зміну тиску всередині сечового міхура за його наповнення.

Мал. 2. А. Волокно скелетного м'яза, клітина серцевого м'яза, гладка м'язова клітина. Б. Саркомір скелетного м'яза. В. Будова гладкого м'яза. Г. Механограма скелетного м'яза та м'язи серця.
Гладким м'язам притаманні самі основні властивості, як і поперечнополосатым скелетним м'язам, а й деякі особливі властивості:
- автоматія, тобто. здатність скорочуватися і розслаблятися без зовнішніх подразнень, а й за рахунок збуджень, що у них самих;
- висока чутливість до хімічних подразників;
- виражена пластичність;
- скорочення у відповідь швидке розтягування.
Скорочення та розслаблення гладких м'язів відбувається повільно. Це сприяє настанню перестальтичних та маятникоподібних рухів органів травного тракту, що призводить до переміщення харчової грудки. Тривале скорочення гладких м'язів необхідно у сфінктерах порожнистих органів і перешкоджає виходу вмісту: жовчі у жовчному міхурі, сечі у сечовому міхурі. Скорочення гладких волокон відбувається незалежно від нашого бажання, під впливом внутрішніх, не підпорядкованих свідомості причин.
Поперечно-смугасті м'язи
Поперечно-смугасті м'язирозташовуються на кістках скелета і скороченням надають руху окремі суглоби і все тіло. утворюють тіло, або сому, тому їх ще називають соматичними, а систему, що іннервує, — соматичною нервовою системою.
Завдяки діяльності скелетної мускулатури здійснюється пересування тіла в просторі, різноманітна робота кінцівок, розширення грудної клітки при диханні, рух голови та хребта, жування, міміка обличчя. Налічується понад 400 м'язів. Загальна маса м'язів становить 40% ваги. Зазвичай середня частина м'яза складається з м'язової тканини та утворює черевце. Кінці м'язів - сухожилля побудовані із щільної сполучної тканини; вони з'єднуються з кістками за допомогою окістя, але можуть прикріплюватися і до іншого м'яза, і до сполучного шару шкіри. У м'язі м'язові та сухожильні волокна поєднуються в пучки за допомогою пухкої сполучної тканини. Між пучками розташовуються нерви та кровоносні судини. пропорційна кількості волокон, що становлять черевце м'яза.

Мал. 3. Функції м'язової тканини
Деякі м'язи проходять тільки через один суглоб і при скороченні рухають його — односуглобові м'язи. Інші м'язи проходять через два або кілька суглобів - багатосуглобові, вони роблять рух у кількох суглобах.
Наприкінці м'язи, прикріплені до кісток, наближаються друг до друга, а розміри м'язи (довжина) зменшується. Кістки, з'єднані суглобами, діють як важелі.
Змінюючи положення кісткових важелів, м'язи діють суглоби. При цьому кожен м'яз впливає на суглоб лише в одному напрямку. У одновісного суглоба (циліндричний, блоковидний) є два м'язи, що діють на нього, або групи м'язів, які є антагоністами: один м'яз — згинач, інший — розгинач. У той самий час кожний суглоб в одному напрямі діє, зазвичай, два м'язи і більше, є синергістами (синергізм — спільну дію).
У двовісного суглоба (еліпсоїдний, м'язовий, сідлоподібний) м'язи групуються відповідно двом його осям, навколо яких відбуваються рухи. До кулястого суглоба, що має три осі руху (багатовісний суглоб), м'язи належать з усіх боків. Так, наприклад, у плечовому суглобі є м'язи-згиначі та розгиначі (рухи навколо фронтальної осі), що відводять та приводять (сагітальна вісь) і обертачі навколо поздовжньої осі, всередину та назовні. Розрізняють три види роботи м'язів: долає, що поступається і утримує.
Якщо скорочення м'язи змінюється становище частини тіла, то долається сила опору, тобто. виконується долаюча робота. Робота, при якій сила м'яза поступається дії сили тяжіння і вантажу, що утримується, називається поступається. У цьому випадку м'яз функціонує, проте він не коротшає, а подовжується, наприклад, коли неможливо підняти або утримати на вазі тіло, що має велику масу. При великому зусиллі м'язів доводиться опустити це тіло на якусь поверхню.
Утримуюча робота виконується завдяки скороченню м'язів, тіло або вантаж утримується в певному положенні без переміщення в просторі, наприклад, людина тримає вантаж, не рухаючись. У цьому м'язи скорочуються без зміни довжини. Сила скорочення м'язів врівноважує масу тіла та вантажу.
Коли м'яз, скорочуючись, перемішує тіло або його частини в просторі, вони виконують долає або поступається роботу, яка є динамічною. Статистичною є робота, що утримує, при якій не відбувається рухів всього тіла або його частини. Режим, при якому м'яз може вільно коротшати, називається ізотонічним(Не відбувається зміни напруги м'яза і змінюється тільки його довжина). Режим, при якому м'яз не може укоротитися, називається ізометричним- Змінюється тільки напруга м'язових волокон.

Мал. 4. М'язи людини
Будова поперечно-смугастих м'язів
Скелетні м'язи складаються з великої кількості м'язових волокон, які поєднуються в м'язові пучки.
В одному пучку міститься 20-60 волокон. М'язові волокна є клітинами циліндричної форми довжиною 10-12 см і діаметром 10-100 мкм.
Кожне м'язове волокно має оболонку (сарколему) та цитоплазму (саркоплазму). У саркоплазмі знаходяться всі компоненти тваринної клітини і вздовж осі м'язового волокна розташовуються тонкі нитки. міофібрили,Кожна міофібрилла складається з протофібрил,до складу яких включені нитки білків міозину і актину, що є скоротливим апаратом м'язового волокна. Міофібрили розділені між собою перегородками, які називаються Z-мембранами, на ділянки - саркомери.На обох кінцях саркомерів до Z-мембрані прикріплені тонкі актинові нитки, а в середині розташовані товсті нитки міозину. Нитки актина своїми кінцями частково входять між міозиновими нитками. У світловому мікроскопі нитки міозину виглядають у вигляді світлої смужки темному диску. При електронній мікроскопії скелетні м'язи виглядають смугастими (поперечно-смугастими).

Мал. 5. Поперечні містки: Ак - актин; Мз - міозин; Гол - головка; Ш - шия
На бокових сторонах міозинової нитки є виступи, що отримали назву поперечних містків(рис. 5), які розташовані під кутом 120° стосовно осі міозинової нитки. Актинові філаменти виглядають як подвійний нитки, закрученої в подвійну спіраль. У поздовжніх борозенках актинової спіралі є нитки білка тропоміозину, до яких приєднаний білок тропонін. У стані спокою молекули білка тропоміозину розташовані таким чином, щоб запобігати прикріпленню поперечних містків міозину до актинових ниток.

Мал. 6. А — організація циліндричних волокон у кістяковому м'язі, прикріпленому до кісток сухожиллями. Б - структурна організація філаментів у волокні скелетного м'яза, що створює картину поперечних смуг.

Мал. 7. Будова актину та міозину
У багатьох місцях поверхнева мембрана поглиблюється у вигляді мікротрубок всередину волокна, перпендикулярно до його поздовжньої осі, утворюючи систему поперечних трубочок(Т-система). Паралельно міофібрилам і перпендикулярно поперечним трубочкам між міофібрил розташована система поздовжніх трубочок(Саркоплазматичний ретикулум). Кінцеві розширення цих трубочок - термінальні цистерни -підходять дуже близько до поперечних трубочок, утворюючи разом із ними звані тріади. У цистернах зосереджено основну кількість внутрішньоклітинного кальцію.
Механізм скорочення скелетного м'яза
М'язова тканинаскладається з клітин, званих м'язовими волокнами. Зовні волокно оточене оболонкою - сарколемою. Усередині сарколеми міститься цитоплазма (саркоплазма), що містить ядра та мітохондрії. У ній міститься безліч скорочувальних елементів, званих міофібрилами. Міофібрили проходять від одного кінця м'язового волокна до іншого. Вони існують порівняно короткий термін — близько 30 діб, після чого й відбувається їхня повна зміна. У м'язах йде інтенсивний синтез білка, необхідний освіти нових міофібрил.
М'язове волокномістить велику кількість ядер, які розташовуються безпосередньо під сарколеммою, оскільки основна частина м'язового волокна зайнята міофібрилами. Саме наявність великої кількості ядер забезпечує синтез нових міофібрил. Така швидка зміна міофібрилу забезпечує високу надійність фізіологічних функцій м'язової тканини.

Мал. 7. А - схема організації саркоплазматичного ретикулума, поперечних трубочок і міофібрил. Б - схема анатомічної структури поперечних трубочок та саркоплазматичного ретикулуму в індивідуальному волокні скелетного м'яза. В - роль саркоплазматичного ретикулуму в механізмі скорочення скелетного м'яза
Кожна міофібрилла складається з правильно чергуються світлих і темних ділянок. Ці ділянки, маючи різні оптичні властивості, створюють поперечну смугастість м'язової тканини.
У скелетному м'язі скорочення викликається надходженням до неї імпульсу нервом. Передача нервового імпульсу з нерва на м'яз здійснюється через нервово-м'язовий синапс (контакт).
Поодинокий нервовий імпульс, або одноразове подразнення, призводить до елементарного скорочувального акту - поодинокого скорочення. Початок скорочення не збігається з моментом нанесення роздратування, оскільки існує прихований, або латентний період (інтервал між нанесенням роздратування і початком скорочення м'яза). У цей період відбувається розвиток потенціалу дії, активація ферментних процесів та розпад АТФ. Після цього починається скорочення. Розпад АТФ у м'язі призводить до перетворення хімічної енергії на механічну. Енергетичні процеси завжди супроводжуються виділенням тепла і теплова енергія зазвичай є проміжною між хімічною та механічними енергіями. У м'язі ж хімічна енергія перетворюється безпосередньо на механічну. Але тепло у м'язі утворюється і за рахунок укорочення м'яза, і під час його розслаблення. Тепло, що утворюється у м'язах, відіграє велику роль у підтримці температури тіла.
На відміну від серцевого м'яза, який має властивість автоматики, тобто. вона здатна скорочуватися під впливом імпульсів, що виникають у ній самій, і на відміну від гладкої мускулатури, також здатної до скорочення без надходження сигналів ззовні, скелетний м'яз скорочується тільки при надходженні до неї сигналів. Безпосередньо сигнали до м'язових волокон надходять за аксонами рухових клітин, які розташовані в передніх рогах сірої речовини спинного мозку (мотонейронів).
Рефлекторний характер діяльності м'язів та координація м'язових скорочень
Скелетні м'язи, на відміну від гладких, здатні здійснювати довільні швидкі скорочення і робити цим значну роботу. Робочим елементом м'яза є м'язове волокно. Типове м'язове волокно є структури з кількома ядрами, відсунутими на периферію масою скорочувальних міофібрил.
М'язові волокна мають три основні властивості:
- збудливістю - здатністю відповідати на дії подразника генерацією потенціалу дії;
- провідністю - здатністю проводити хвилю збудження вздовж усього волокна в обидві сторони від точки подразнення;
- скоротливістю - здатністю скорочуватися або змінювати напругу при збудженні.
У фізіології є поняття рухової одиниці, під якою мається на увазі один руховий нейрон і всі м'язові волокна, які нейрон іннервує. Двигуни одиниці бувають різними за обсягом: від 10 м'язових волокон на одиницю для м'язів, що виконують точні рухи, до 1000 і більше волокон на рухову одиницю для м'язів «силової спрямованості». Характер роботи скелетних м'язів може бути різним: статична робота (підтримка пози, утримання вантажу) та динамічна робота (переміщення тіла чи вантажу у просторі). М'язи беруть участь також у пересуванні крові та лімфи в організмі, виробленні тепла, актах вдиху та видиху, є своєрідними депо для води та солей, захищають внутрішні органи, наприклад м'язи черевної стінки.
Для скелетного м'яза характерні два основні режими скорочення — ізометричний та ізотонічний.
Ізометричний режим проявляється в тому, що в м'язі під час його активності наростає напруга (генерується сила), але через те, що обидва кінці м'язи фіксовані (наприклад, при спробі підняти дуже великий вантаж), вона не вкорочується.
Ізотонічний режим проявляється в тому, що м'яз спочатку розвиває напругу (силу), здатну підняти даний вантаж, а потім м'яз вкорочується - змінює свою довжину, зберігаючи напругу, що дорівнює вазі вантажу, що утримується. Чисто ізометричного чи ізотонічного скорочення практично спостерігати не можна, але існують прийоми так званої ізометричної гімнастики, коли спортсмен напружує м'язи без зміни довжини. Ці вправи більшою мірою розвивають силу м'язів, ніж вправи з ізотонічними елементами.
Скорочувальний апарат скелетного м'яза представлений міофібрилами. Кожна міофібрилла діаметром 1 мкм складається з кількох тисяч протофібрил - тонких, подовжених полімеризованих молекул білків міозину та актину. Міозинові нитки вдвічі тонші за актинові, і в стані спокою м'язового волокна актинові нитки вільними кільцями входять між міозиновими нитками.
У передачі збудження велику роль відіграють іони кальцію, які входять до міжфібрилярного простору і запускають механізм скорочення: взаємне втягування щодо один одного актинових та міозинових ниток. Втягування ниток відбувається за обов'язкової участі АТФ. В активних центрах, розташованих на одному з кінців міозинових ниток, АТФ розщеплюється. Енергія, що виділяється при розщепленні АТФ, перетворюється на рух. У скелетних м'язах запас АТФ невеликий — лише на 10 одиночних скорочень. Тому потрібен постійний синтез АТФ, який іде трьома шляхами: перший — за рахунок запасів креатинфосфату, які обмежені; другий — гліколітичний шлях при анаеробному розщепленні глюкози, коли на одну молекулу глюкози утворюється дві молекули АТФ, але одночасно утворюється молочна кислота, яка гальмує активність гліколітичних ферментів, і нарешті третій — аеробне окислення глюкози і жирних кислот в циклі Кребса, що відбувається в молекулах зи. Останній процес є найбільш економічним, але дуже повільним. Постійне тренування активізує третій шлях окислення, у результаті підвищується витривалість м'язів до тривалим навантаженням.
внутрішніх органів, шкіри, судин.
Скелетні м'язиРазом зі скелетом становлять опорно-рухову систему організму, що забезпечує підтримку пози і рух тіла у просторі. Крім того, вони виконують захисну функцію, оберігаючи внутрішні органи від пошкоджень.
Скелетні м'язи є активною частиною опорно-рухового апарату, що включає також кістки та їх зчленування, зв'язки, сухожилля. Маса м'язів може досягати 50% від загальної маси тіла.
З функціональної точки зору до рухового апарату можна віднести і моторні нейрони, що посилають нервові імпульси до м'язових волокон. Тіла моторних нейронів, що іннервують аксонами скелетну мускулатуру, розташовуються в передніх рогах спинного мозку, а іннервують м'язи щелепно-лицьової області - в моторних ядрах стовбура мозку. Аксон мотонейрона при вході в скелетний м'яз розгалужується, і кожна гілочка бере участь у формуванні нервово-м'язового синапсу на окремому м'язовому волокні (рис. 1).
Мал. 1. Розгалуження аксона моторного нейрона на аксонні термінали. Електронограма

Мал. Будова скелетного м'яза людини
Скелетні м'язи складаються з м'язових волокон, які поєднуються в м'язові пучки. Сукупність м'язових волокон, иннервируемых гілочками аксона одного моторного нейрона, називають рухової (чи моторної) одиницею. У очних м'язах 1 рухова одиниця може містити 3-5 м'язових волокон, у м'язах тулуба - сотні волокон, у камбалоподібному м'язі - 1500-2500 волокон. М'язові волокна 1 рухової одиниці мають однакові морфофункціональні властивості.
функціями скелетних м'язівє:
- пересування тіла у просторі;
- переміщення частин тіла щодо один одного, у тому числі здійснення дихальних рухів, що забезпечують вентиляцію легень;
- підтримання становища та пози тіла.
Скелетні м'язи разом зі скелетом складають опорно-рухову систему організму, яка забезпечує підтримку пози та переміщення тіла у просторі. Поруч із скелетні м'язи і скелет виконують захисну функцію, оберігаючи внутрішні органи від ушкодження.
Крім того, поперечно-смугасті м'язи мають значення у виробленні тепла, що підтримує температурний гомеостаз, і в депонуванні деяких поживних речовин.
Мал. 2. Функції кістякових м'язів
Фізіологічні властивості кістякових м'язів
Скелетні м'язи мають такі фізіологічні властивості.
Збудливість.Забезпечується властивістю плазматичної мембрани (сарколеми) відповідати збудженням надходження нервового імпульсу. Через більшу різницю потенціалу спокою мембрани поперечно-смугастих м'язових волокон (Е 0 близько 90 мВ) збудливість їх нижча, ніж нервових волокон (Е 0 близько 70 мВ). Амплітуда потенціалу дії у них більше (близько 120 мВ), ніж в інших збуджуваних клітин.
Це дозволяє практично досить легко реєструвати біоелектричну активність скелетних миші. Тривалість потенціалу дії становить 3-5 мс, що визначає коротку тривалість фази абсолютної рефрактерності збудженої м'язової мембрани волокон.
Провідність.Забезпечується властивістю плазматичної мембрани формувати локальні кругові струми, генерувати та проводити потенціал дії. В результаті потенціал дії поширюються по мембрані вздовж м'язового волокна і вглиб поперечними трубочками, що формуються мембраною. Швидкість проведення потенціалу дії становить 3-5 м/с.
Короткість.Являє собою специфічну властивість м'язових волокон змінювати свою довжину і напругу за збудженням мембрани. Скоротимість забезпечується спеціалізованими скоротливими білками м'язового волокна.
Скелетні м'язи мають також в'язкоеластичними властивостями, що мають важливе значення для розслаблення м'язів.

Мал. Скелетні м'язи людини
Фізичні властивості скелетних м'язів
Скелетні м'язи характеризуються розтяжністю, еластичністю, силою та здатністю виконувати роботу.
Розтяжність -здатність м'яза змінювати довжину під дією сили, що розтягує.
Еластичність -здатність м'язи відновлювати первісну форму після припинення дії розтягуючої або деформуючої сили.
- здатність м'яза піднімати вантаж. Для порівняння сили різних м'язів визначають їхню питому силу шляхом поділу максимальної маси на число квадратних сантиметрів її фізіологічного перерізу. Сила скелетного м'яза залежить від багатьох факторів. Наприклад, від числа рухових одиниць, що збуджуються на даний момент часу. Також вона залежить від синхронності роботи рухових одиниць. Сила м'яза залежить від вихідної довжини. Існує певна середня довжина, коли м'яз розвиває максимальне скорочення.
Сила гладких м'язів також залежить від вихідної довжини, синхронності збудження м'язового комплексу, і навіть від концентрації іонів кальцію всередині клітини.
Здатність м'яза виконувати роботу.Робота м'яза визначається добутком маси піднятого вантажу на висоту підйому.
Робота м'язів зростаєте збільшенням маси вантажу, що піднімається, але до певної межі, після якого збільшення вантажу призводить до зменшення роботи, тобто. знижується висота підйому. Максимальна робота здійснюється м'язом при середніх навантаженнях. Це називається законом середніх навантажень. Розмір м'язової роботи залежить від кількості м'язових волокон. Чим товщі м'яз, тим більший вантаж може підняти. Тривала напруга м'яза призводить до її втоми. Це зумовлено виснаженням енергетичних запасів у м'язі (АТФ, глікоген, глюкоза), накопиченням молочної кислоти та інших метаболітів.
Допоміжні властивості кістякової мускулатури
Розтяжність - це здатність м'яза змінювати свою довжину під дією сили, що розтягує її. Еластичність - здатність м'язи приймати свою початкову довжину після припинення дії сили, що розтягує або деформує. Живий м'яз має малу, але досконалу еластичність: вже невелика сила здатна викликати відносно велике подовження м'яза, а повернення її до початкових розмірів є повним. Ця властивість дуже важлива для здійснення нормальних функцій скелетних м'язів.
Сила м'яза визначається максимальним вантажем, який м'яз може підняти. Для порівняння сили різних м'язів визначають їхню питому силу, тобто. максимальний вантаж, який м'яз може підняти, ділять на число квадратних сантиметрів її фізіологічного поперечного перерізу.
Здатність м'яза виконувати роботу.Робота м'яза визначається добутком величини піднятого вантажу на висоту підйому. Робота м'яза поступово збільшується зі збільшенням вантажу, але до певної межі, після якого збільшення вантажу призводить до зменшення роботи, оскільки знижується висота підйому вантажу. Отже, максимальна робота м'язом проводиться за середніх величин навантажень.
Втома м'язів.М'язи не можуть працювати безперервно. Тривала робота призводить до зниження їхньої працездатності. Тимчасове зниження працездатності м'яза, що настає при тривалій роботі та зникає після відпочинку, називається втомою м'яза. Прийнято розрізняти два види втоми м'язів: хибне та дійсне. При хибному стомленні втомлюється не м'яз, а особливий механізм передачі імпульсів з нерва на м'яз, званий синапсом. У синапсі виснажуються резерви медіаторів. При справжньому стомленні в м'язі відбуваються такі процеси: накопичення недоокислених продуктів розпаду поживних речовин внаслідок недостатнього надходження кисню, виснаження запасів джерел енергії, необхідної для м'язового скорочення. Втома проявляється зменшенням сили скорочення м'яза та ступеня розслаблення м'яза. Якщо м'яз на деякий час припиняє роботу і перебуває у стані спокою, то відновлюється робота синапсу, а з кров'ю видаляються продукти обміну та доставляються поживні речовини. Таким чином, м'яз знову набуває здатності скорочуватися і виконувати роботу.
Поодиноке скорочення
Роздратування м'яза або рухового нерва, що його іннервує, одиночним стимулом викликає одиночне скорочення м'яза. Розрізняють три основні фази такого скорочення: латентна фаза, фаза укорочення та фаза розслаблення.
Амплітуда одиночного скорочення ізольованого м'язового волокна від сили роздратування залежить, тобто. підпорядковується закону «усі чи нічого». Однак скорочення цілого м'яза, що складається з безлічі волокон, при її прямому подразненні залежить від сили подразнення. При граничній силі струму в реакцію залучається лише невелика кількість волокон, тому скорочення м'яза ледве помітно. Зі збільшенням сили роздратування кількість волокон, охоплених збудженням, зростає; скорочення посилюється до того часу, поки всі волокна не виявляються скороченими («максимальне скорочення») — цей ефект називається сходами Боудича. Подальше посилення подразнюючого струму скорочення м'язи не впливає.

Мал. 3. Поодиноке скорочення м'яза: А - момент подразнення м'яза; а-6 - прихований період; 6-в - скорочення (укорочення); в-г - розслаблення; г-д - послідовні еластичні коливання.
Тетанус м'язи
У природних умовах до скелетного м'яза з центральної нервової системи надходять не поодинокі імпульси збудження, які є для неї адекватними подразниками, а серії імпульсів, на які м'яз відповідає тривалим скороченням. Тривале скорочення м'яза, що виникає у відповідь на ритмічне подразнення, отримало назву тетанічного скорочення, або тетануса. Розрізняють два види тетанусу: зубчастий і гладкий (рис. 4).
Гладкий тетанусвиникає, коли кожен наступний імпульс збудження надходить у фазу укорочення, а зубчастий -у фазу розслаблення.
Амплітуда тетанічного скорочення перевищує амплітуду одиночного скорочення. Академік Н.Є. Введенський обґрунтував мінливість амплітуди тетануса неоднаковою величиною збудливості м'яза і ввів у фізіологію поняття оптимуму та песимуму частоти подразнення.
Оптимальноюназивається така частота подразнення, при якій кожне наступне подразнення надходить у фазу підвищеної збудливості м'яза. У цьому розвивається тетанус максимальної величини (оптимальний).
Песимальноїназивається така частота подразнення, за якої кожне наступне подразнення здійснюється у фазу зниженої збудливості м'яза. Величина тетанусу при цьому буде мінімальною (песимальною).

Мал. 4. Скорочення скелетного м'яза при різній частоті подразнення: I - скорочення м'яза; II - Позначка частоти подразнення; а - поодинокі скорочення; б-зубчастий тетанус; в - гладкий тетанус
Режими м'язових скорочень
Для скелетних м'язів характерні ізотонічний, ізометричний та змішаний режими скорочення.
При ізотонічномускорочення м'яза змінюється її довжина, а напруга залишається постійною. Таке скорочення відбувається у тому випадку, коли м'яз не долає опору (наприклад, не переміщує вантаж). У природних умовах близькими до ізотонічного типу скорочення є скорочення м'язів язика.
При ізометричномускорочення у м'язі під час її активності наростає напруга, але через те, що обидва кінці м'язи фіксовані (наприклад, м'яз намагається підняти великий вантаж), він не коротшає. Довжина м'язових волокон залишається постійною, змінюється лише ступінь їхньої напруги.
Скорочуються за аналогічними механізмами.
В організмі скорочення м'язів ніколи не бувають чисто ізотонічними чи ізометричними. Вони мають змішаний характер, тобто. відбувається одночасна зміна і довжини, і напруження м'яза. Такий режим скорочення називається аукстонічним,якщо переважає напруга м'яза, або ауксометричним,якщо переважає скорочення.
Міністерство охорони здоров'я Республіки Білорусь
УО «Гомельський державний медичний університет»
Обговорено на засіданні кафедри
Протокол № 200 року
із загальної гістології
для студентів лікувального, медико-діагностичного та
медико-профілактичного факультетів
Тема: «М'язові тканини»
Час – 90 хвилин
Навчальні та виховні цілі:
1. Викласти загальні принципи морфологічної організації та функціонування м'язових тканин.
2. Ознайомити з класифікацією м'язових тканин та викласти їх структурно-функціональні особливості та локалізацію.
3. Дати уявлення про молекулярні та клітинні механізми м'язового скорочення.
ЛІТЕРАТУРА
1. Гістологія, цитологія та ембріологія,
(За ред. Ю. І. Афанасьєва). - М.: Медицина, 1999.
2. Биков В.Л. - приватна гістологія людини. - С.-П.: Sotis, 2000.
3. Гістологія (під ред. Е. Г. Улумбекова та Ю. А. Челишева) - М.: ГОЕТАР, 1997.
4. Гістологія у питаннях та відповідях (за ред. Слуки Б.А.) – Мозир: Білий вітер, 2000.
5. Мяделець О. Д. - Курс лекцій з приватної гістології - Вітебськ, 1996.
МАТЕРІАЛЬНЕ ЗАБЕЗПЕЧЕННЯ
1. Мультимедійна презентація
РОЗРАХУНОК НАВЧАЛЬНОГО ЧАСУ
| № | Перелік навчальних питань | Продовж |
| 5 хв. | ||
| Організація м'язового волокна. | 10 хв. | |
| Механізм м'язового скорочення | 5 хв. | |
| Апарат передачі збудження | 5 хв. | |
| Серцева м'язова тканина. | 10 хв. | |
| Загальна характеристика робітничих кардіоміоцитів. | 5 хв. | |
| Провідна система серця. | 10 хв. | |
| Секреторні кардіоміоцити. | 5 хв | |
| Гладка м'язова тканина | 10 хв. | |
| Гістогенез та регенерація м'язових тканин. | 5 хв. |
Усього 90 хвилин
Питання 1.Загальні структурні особливості м'язових тканин.
Поєднує кілька різних видів, але основна властивість загальна – скоротливість. Тому всі м'язові тканини мають подібні структурні особливості:
1. Клітини витягнутої форми та об'єднані у тяжі, або навіть у симпласти (м'язові волокна).
2. Цитоплазма заповнена міофіламентами – нитками із скорочувальних білків (міозин та актин), взаємне ковзання яких забезпечує скорочення. Характер розташування міофіламентів залежить від виду м'язової тканини.
3. Високі енергетичні запити вимагають безлічі мітохондрій, включень міоглобіну, жиру та глікогену.
4. Гладка ЕПС спеціалізована на накопиченні Сa 2+, який ініціює скорочення.
Відповідно до морфо-функціональної класифікації виділяють:
1. Поперечно-смугасті м'язові тканини. У тому цитоплазмі головний компонент – міофібрили (органели загального значення), що й створюють ефект смугастість. Цих тканин два види:
Скелетна. Утворюється із міотомів сомітів.
Серцева. Утворюється з вісцерального листка спланхнотома.
2. Гладка м'язова тканина. Її клітини не містять міофібрил. Утворюється із мезенхіми.
До цієї ж групи відносять міоепітеліальні клітини, які мають ектодермальне походження і м'язи райдужної оболонки ока, які мають нейральне походження.
Скелетна м'язова тканина.
Питання 2. Організація м'язового волокна.
Структурно-функціональною одиницею цієї тканини є м'язове волокно. Це довгий цитоплазматичний тяж з безліччю ядер, які лежать одразу під плазмолемою. М'язове волокно в ембріогенезі утворюється при злитті клітин – міобластів, тобто є клітинним похідним – симпласт.
М'язове волокно зберігає загальний план клітинної організації. У ньому є всі органели загального значення, багато включень, і навіть органели спеціального значення. Усі компоненти волокна адаптовані до виконання головної функції – скорочення – і поділяються кілька апаратів.
Скоротливий апарат складається з міофібрил. Це органели, які тягнуться вздовж усього волокна і займають більшу частину всього об'єму цитоплазми. Вони можуть значно змінювати свою довжину.
Апарат білкового синтезу представлений, в основному, вільними рибосомами та спеціалізований на виробленні білків для побудови міофібрил.
Апарат передачі збудження утворений саркотубулярною системою. Вона включає гладку ЕПС та Т-трубочки. Гладка ЕПС (саркоплазматична мережа) має вигляд плоских цистерн, які обплітають усі міофібрили. Вона служить для накопичення Сa 2+. Її мембрани здатні швидко випускати кальцій назовні, що необхідно для укорочення міофібрилу, а потім активно закачує його всередину. Зовнішня мембрана м'язового волокна (сарколемма) утворює численні трубчасті вп'ячування, що пронизують все волокно в поперечних напрямках. Їхня сукупність називають Т-системою. Т-трубочки тісно контактують з мембранами ЕПС, утворюючи єдину саркотубулярну систему. До кожної Т-трубочки …..
Енергетичний апарат складений мітохондріями та включеннями. Мітохондрії великі витягнуті і лежать, переважно ланцюжками, заповнюючи весь простір між міофібрилами. Субстратами для отримання АТФ є глікоген і ліпідні краплі. Включення міоглобіну – специфічного м'язового пігменту, що забезпечують волокна киснем у разі тривалої та напруженої роботи м'язів.
Лізосомальний апарат розвинений слабо. Служить головним чином для процесів внутрішньоклітинної регенерації.
Питання 3 Механізм м'язового скорочення.Для його розуміння необхідно ознайомитись із молекулярною організацією міофібрил – органел, спеціалізованих на скороченні. Це довгі тяжі, що утворюють поздовжні пучки по тисячі і більше міофібрил, майже повністю заповнюють цитоплазму волокна.
Кожна міофібрилла побудована з величезної кількості актинових та міозинових філаментів.
Тонкі актинові нитки побудовані з глобулярних молекул білка актину, які поєднуються у два спірально закручені ланцюжки. Товстіша міозинова нитка побудована з 300-400 молекул білка міозину. Кожна молекула включає довгий хвіст, якого з одного краю прикріплена рухлива головка. Головки можуть міняти кут свого нахилу. Хвости безлічі молекул укладаються щільним пучком, формуючи стрижень філаменту. Головки залишаються на поверхні. На двох краях нитки головки лежать різноспрямовано.
Завдяки додатковим білкам, міофіламенти мають стабільний діаметр та стабільну довжину близько 1 мкм. Філаменти одного виду утворюють акуратно підігнані пучки або стоси. Міофібрили утворені з багаторазово чергуються пучків актинових і міозинових ниток. Висока впорядкованість розташування міофіламентів досягається за допомогою різних білків цитоскелета. Наприклад, білок актинін формує Z-лінію (телофрагму), до якої з обох боків приєднуються краї тонких актинових ниток. Так утворюється актинова стопка. Інші білки організують у стос товсті міозинові нитки, прошнуровуючи їх у середині. Вони утворюють М-лінію (мезофрагму). При чергуванні стопок вільні кінці тонких і товстих ниток заходять один за одного, забезпечуючи взаємне ковзання щодо одного в момент скорочення. В результаті такої організації в міофібрилі багаторазово повторюються світлі ділянки, які називаються I-дисками (ізотропні), і темні ділянки, які називаються А-дисками (анізотропні). Це створює ефект поперечної смугастість. Ізотропні ділянки відповідають центральній частині актинового стосу і містять лише тонкі нитки. Анізотропні диски відповідають цілій міозиновій стопці, і в них входять суто міозинова частина (Н-смужка) і ті ділянки, де кінці тонких і товстих ниток перекриваються.
Ділянку між двома Z-лініями називають саркомером. Саркомір є структурною одиницею міофібрили. (20 тисяч саркомірів на міофібрилу). Сувора організація міофібрил забезпечується широким набором різних білків цитоскелета.
При скороченні довжина міофібрили зменшується за рахунок одночасного скорочення всіх I-дисків. Довжина кожного саркомера зменшується в 1,5-2 рази. Процес скорочення пояснюється теорією ковзання Хакслі, згідно з якою в момент скорочення вільні кінці актинових і міозинових ниток вступають в молекулярні взаємодії і глибше засуваються один щодо одного. Ковзання починається з того, що виступаючі міозинові головки утворюють містки з активними центрами актинових філаментів. Потім кут нахилу головки зменшується, містки здійснюють хіба що гребкові рухи, зміщуючи і актинову нитку. Після цього місток розмикається, що супроводжується гідролізом молекули 1 АТФ. Зв'язування міозинових головок з актиновою ниткою регулюється спеціальними білками. Це тропонін та тропоміозин, які вбудовані в актинову нитку, та перешкоджають контакту з міозиновими головками. При підвищенні в гіалоплазмі концентрації Са 2+ відбувається зміна конформаційного стану цих регуляторних білків та їхня блокуюча дія знімається. Гребкові рухи повторюються сотні разів за одне м'язове скорочення. Розслаблення настає лише після того, як знизиться концентрація Ca 2+ .
Запитання 4. Апарат передачі збудження.
Скорочення запускається нервовим імпульсом, який через моторну бляшку передається на мембрану м'язового волокна, викликаючи хвилю деполяризації, яка миттєво охоплює Т-трубочки. Вони тягнуться від поверхні крізь усі волокна, по дорозі кільцями оточуючи міофібрили. Порожнини гладкої ЕПС, заповнені кальцієм, чохлом обплітають міофібрили, тісно контактуючи з Т-трубочками. З двох сторін до кожної Т-трубочки належать великі мембранні порожнини ЕПС (термінальні цистерни). Такий комплекс називають тріадою. На кожен саркомір припадає дві тріади. Завдяки мембранним контактам деполяризація Т-трубочок змінює стан мембранних білків ЕПС, що призводить до відкриття кальцієвих каналів та виходу кальцію в гіалоплазму. Відбувається скорочення. Тріади сполучають процеси збудження та скорочення. Після викиду спеціальні мембранні насоси активно закачують Ca 2+ назад в ЕПС, де він з'єднується із Са-зв'язуючим білком.
Запитання 5. Серцева м'язова тканинаутворює м'язову стінку серця – міокард. Її морфо-функціональна одиниця – окрема клітина – кардіоміоцит. Клітини з'єднані один з одним особливими структурами - вставними дисками, і в результаті утворюється тривимірна мережа з клітинних тяжів (функціональний синцит), що забезпечує синхронність скорочення під час систоли.
Кардіоміоцити – витягнуті клітини з кількома відгалуженнями, покриті поверх плазмолеми базальною мембраною. Їхні ядра (1 або 2) лежать центрально.
У складі міокарда виділяють кілька популяцій кардіоміоцитів:
А) скорочувальні чи робітники
Б) провідні
В) секреторні
Запитання 6. Робочі кардіоміоцитискладають основну масу міокарда та забезпечують скорочення. Їхня організація подібна до м'язових волокон, але має ряд відмінностей.
Скоротливий апарат. Міофібрили в цілому мають подовжній напрямок, але багато разів анастомозують один з одним.
Саркотубулярна мережа розвинена слабше. Т-трубочки ширші, лежать рідше і кожна контактує лише з однією цистерною ЕПС (діада). При збудженні частина Ca 2+ надходить у гіалоплазму з міжклітинного простору через плазмолемму та мембрани Т-трубочок і лише після цього відбувається Са-індукований викид Ca 2+ з ЕПС.
Енергетичний апарат. Мітохондрії багато, вони великі із щільно упакованими христами, оскільки енергетичні запити міокарда дуже високі. Між собою вони об'єднані особливими сполуками – міжмітохондріальними контактами та утворюють єдину функціональну систему – мітохондріон. Така інтеграція є виключно важливою для швидкого та синхронного скорочення міокарда. Субстрати для отримання АТФ поставляються ліпідними краплями, включеннями глікогену та міоглобіну. Самі мотохондрії здатні накопичувати кальцій.
Кінці сусідніх клітин або їх відгалуження, що стикуються, з'єднуються вставними дисками. Диск має східчасту форму. Поперечні ділянки утворені десмосомами та надають з'єднанню механічної міцності. Поздовжні ділянки містять безліч щілинних контактів - нексусів, яких особливо багато в передсердях. Завдяки іонним каналам нексусів збудження швидко поширюється вздовж усього м'яза.
Міокард рясно кровопостачається. Усі проміжки між кардіоміоцитами заповнені пухкою сполучною тканиною, в якій розгалужуються капіляри. Тут же закінчуються розгалуження нервових волокон вегетативної нервової системи. На відміну від скелетної м'язової тканини тут утворюються не нейром'язові синапси (моторні бляшки), а лише варикозні розширення. На скорочувальну активність кардіоміоцитів нервова система має лише регуляторний вплив. Вегетативна система лише збільшує (симпатичний відділ) або зменшує (парасимпатичний відділ) частоту та силу серцевих скорочень.
Ритмічна генерація імпульсів, що змушують серце постійно скорочуватися, забезпечується спеціальними клітинами самого міокарда. Сукупність цих клітин називається провідною системою серця, а здатність серця скорочуватися незалежно від нервових стимулів – автоматією серця.
Питання 7. Провідна системавключає спеціалізовані кардіомміоцити, які називаються також атиповими. До них відносять:
Пейсмекерні клітини чи водії ритму. Їхня головна властивість – нестійкий потенціал спокою зовнішньої мембрани. Завдяки К/Na насосу натрію завжди більше всередині клітини, а калію зовні. Ця різниця іонів і створює електричний потенціал з обох боків плазмолеми. При певній стимуляції у мембрані відкриваються натрієві канали, натрій спрямовується назовні та мембрана деполяризується. У пейсмекерних клітин завдяки постійному невеликому витоку іонів плазмолема регулярно деполяризується без зовнішніх сигналів. Це спричиняє потенціал дії, що поширюється і на сусідні клітини, викликаючи їхнє скорочення. Головні водії ритму – це кардіоміоцити синусно-передсердного вузла. Щохвилини вони генерують 60-90 імпульсів. Водії ритму другого порядку утворюють передсердно-шлуночковий вузол. Вони генерують імпульси з частотою 40 імпульсів на хвилину, і в нормі їхня активність пригнічується головними пейсмекерами. Пейсмекерні кардіоміоцити – дрібні світлі клітини із великим ядром. Їхній скорочувальний апарат розвинений слабо.
Провідні кардіоміоцити забезпечують швидку передачу збудження від водіїв ритму до робочих кардіоміоцитів. Ці клітини об'єднані в довгі тяжі, що формують пучок Гіса та волокна Пуркіньє. Пучок Гіса складений клітинами середнього розміру з рідкими довгими звивистими міофібрилами та дрібними мітохондріями. Волокна Пуркіньє містять найбільші кардіоміоцити, які можуть контактувати відразу з кількома робочими клітинами. Міофібрили тут утворюють рідкісну невпорядковану мережу, Т-система не розвинена. Вставних дисків немає, але клітини об'єднані безліччю нексусів, що забезпечує високу швидкість проведення імпульсів.
Запитання 8. Секреторні кардіоміоцити. У передсердях зустрічаються відросткові клітини, в яких добре розвинена грЕПС, комплекс Гольджі та містяться секреторні гранули. Міофібрили розвинені дуже слабо, оскільки основною функцією є вироблення гормону (натрійуретичний фактор), що регулює обмін електролітів та артеріальний тиск.
Питання 9. Гладка м'язова тканина.
Збудована з гладких міоцитів. Скоротливі філаменти у цих клітинах немає жорсткої впорядкованості і миофибриллы у яких не утворюються. Внаслідок цього відсутня і поперечна смугастість. Гладкі міоцити досить великі клітини веретеноподібної форми, покриті зверху базальною мембраною, яка з'єднана з міжклітинною речовиною. У центрі витягнуте ядро, біля полюсів грЕПС, комплекс Гольджі та рибосоми. Клітини секретують компоненти міжклітинної речовини для своєї зовнішньої оболонки, а також деякі ростові фактори та цитокіни. Багато дрібних мітохондрій. Саркоплазматична мережа (гладка ЕПС) розвинена слабо, виконує роль кальцієвого депо. Системи Т-трубочок немає, і їхню функцію виконують кавеоли. Кавеоли - це дрібні вп'ячування плазмолеми у вигляді бульбашок. Вони містять високі концентрації кальцію, що захоплюють з міжклітинного простору. У момент збудження Ca 2+ з кавеол виходить назовні, що ініціює звільнення Ca 2+ із саркоплазматичної мережі.
Організація та функціонування скорочувального апарату своєрідні. Актинові та міозинові філаменти дуже численні, але не утворюють міофібрил. Для їх упорядкування у міоциті існує система щільних тілець. Це округлі опорні утворення з білка a-актиніна та десміну. Вони одним кінцем закріплено по 10-20 тонких актинових філаментів. Одні тільця утворюють прикріплювальні пластинки в сарколеммі, інші ланцюжками лежать у гіалоплазмі. Так у міоциті формується стабільна мережа з актинових ниток. Товсті міозиновини нитки мають різну довжину і дуже лабільні.
Кожному скорочення передує викид кальцію, який пов'язується з особливим білком – кальмодуліном. Це активує фермент, що забезпечує швидке збирання міозинових філаментів. Вони вбудовуються між актиновими нитками, утворюють із нею містки, та його голівки починають здійснювати гребкові руху. При взаємному ковзанні ниток щільні тільця зближуються, а клітина загалом коротшає. Таким чином у гладких міоцитах кальцій взаємодіє з міозиновими нитками, а не з актиновими, як у поперечно-смугастих. АТФ-азна активність міозину набагато нижча. Разом із постійним складанням і розбиранням міозинових філаментів це призводить до того, що гладком'язові клітини скорочуються повільніше, але можуть тривалий час підтримувати цей стан (тонічні скорочення). Між собою клітини об'єднані РВСТ, яка вплітається в їх базальні мембрани, а також різними міжклітинними контактами, у тому числі нексусами. Скоротлива активність міоцитів перебуває під контролем нервових та гуморальних факторів. У сполучнотканинних прошарках розташовані варикозні розширення аксонів вегетативної нервової системи. Їхні медіатори деполяризують найближчі міоцити, а до решти збудження передається за щілинними контактами.
Завдяки широкому набору мембранних рецепторів гладкі міоцити чутливі до багатьох біологічно активних речовин (адреналін, гістамін тощо) та реагують по-різному, залежно від органної специфічності.
Запитання 10.Гістогенез та регенерація.
Скелетна м'язова тканина. З міотома сомітів диференціюються одноядерні клітини, що активно діляться - міобласти. Вони зливаються в ланцюжки - м'язові трубочки, численні ядра яких не діляться. У трубочках починається активний синтез скорочувальних білків та формування міофібрил, які поступово заповнюють всю цитоплазму, відтісняючи ядра на периферію. Утворюється м'язове волокно, всередині якого міофібрили постійно оновлюються. Між плазмолемою і базальною мембраною, що її покриває, де-не-де зберігаються одноядерні клітини - міосаттеліти - камбіальні клітини, які можуть ділитися і включати свої ядра до складу волокон. Зростання м'язової тканини у дорослої людини відбувається, переважно за рахунок гіпертрофії волокон, які число залишається постійним. Після пошкодження міосаттеліти можуть зливатись, утворюючи нові волокна.
Серцева м'язова тканина утворюється з міоепікардіальної платівки у складі вісцерального листка спланхнотома. Розподіл кардіоміоцитів закінчується незабаром після народження, але в наступні 10 років можуть формуватися поліплоїдні та двоядерні клітини. Оскільки камбіальних клітин немає, то можлива лише внутрішньоклітинна регенерація та гіпертрофія кардіоміоцитів. Вона відбувається в результаті тривалих фізичних навантажень або в патологічних станах (гіпертонія, вади серця і т.д.). Після загибелі міоцитів (інфркт міокарда) формується сполучнотканинний рубець. Останнім часом встановлено, що окремі передсердні міоцити зберігають здатність до мітозів.
Гладком'язова тканина регенерує як за рахунок гіпертрофії, так і за рахунок гіперплазії.
Гомельський державний медичний інститут
Кафедра гістології, цитології та ембріології
Обговорено на засіданні кафедри
Протокол №
із загальної гістології
для факультету з підготовки іноземних студентів
Тема: «М'язові тканини»
Час – 90 хвилин
Усі м'язові тканини мають однакові структурні особливості:
1. Клітини витягнутої форми.
2. Цитоплазма заповнена міофіламентами – нитками із скорочувальних білків (міозин та актин).
3. Багато мітохондрій, включень міоглобіну, жиру та глікогену (джерела енергії)
4. Гладка ЕПС накопичує Сa 2+ , який необхідний скорочення.
5. Плазмолема м'язових клітин має збудливість.
Морфо-функціональна класифікація:
1. Поперечно-смугасті м'язові тканини. .
Їхні клітини містять міофібрили (органели загального значення), цих тканин два види:
- скелетна . (Утворюється з міотомів сомітів).
- серцева . (Утворюється з вісцерального листка спланхнотома).
2. Гладка м'язова тканина . Її клітини не містять міофібрил. Утворюється із мезенхіми.