დნმ-ის რეპლიკაცია- ეს არის მშობლის დნმ-ის მოლეკულების გაორმაგების პროცესი ცოცხალი ორგანიზმების უჯრედების რეპროდუქციის დროს. ანუ რეპლიკაციის პროცესი წინ უსწრებს უჯრედების გაყოფას. რეპლიკაცია, ისევე როგორც ტრანსკრიფცია და თარგმანი, არის მატრიცული პროცესი. ჯაჭვის რეპლიკაციის დროს დნმ-ის მოლეკულები გამოიყოფა და თითოეული მათგანი ხდება შაბლონი, რომელზედაც სინთეზირებულია ახალი დამატებითი ჯაჭვი. ამ შემთხვევაში, ახალი ჯაჭვების ნუკლეოტიდები წყვილდება შემავსებელიძველი ჯაჭვების ნუკლეოტიდებით (A T-ით, G C-ით). შედეგად, წარმოიქმნება ორი ქალიშვილი ორჯაჭვიანი დნმ-ის მოლეკულა, რომლებიც არ განსხვავდება მშობლის მოლეკულისგან. თითოეული დნმ-ის მოლეკულა შედგება ორიგინალური მშობელი მოლეკულის ერთი ჯაჭვისა და ახლად სინთეზირებული ჯაჭვისგან. კოპირების ამ მექანიზმს ე.წ ნახევრად კონსერვატიული. ყოველი ახლად სინთეზირებული ჯაჭვი ანტიპარალელურიმშობელი. ერთი ძაფის (წინამძღვრის) სინთეზი მუდმივად ხდება, ხოლო მეორე (ჩამორჩენილი) პულსირებულია. ასეთ მექანიზმს ე.წ ნახევრად უწყვეტი.
რეპლიკაციის ჩანგლის სტრუქტურა. წამყვანი ძაფები, ჩამორჩენილი ძაფები, ოკაზაკის ფრაგმენტები. იხილეთ სურათი.
ძირითადი ფერმენტები, რომლებიც მონაწილეობენ დნმ-ის სინთეზში.
დნმ პოლიმერაზების ზოგადი სტრუქტურული მახასიათებლები.
ისინი მუშაობენ იმავე პრინციპით: აგრძელებენ დნმ-ის ჯაჭვს 1 ნუკლეოტიდის დამატებით 3' ბოლოზე. არჩევანი ნაკარნახევია შაბლონის დნმ-ის კომპლემენტარობის მოთხოვნებით. Მახასიათებლები:
რამდენიმე დამოუკიდებელი დომენი, კატა. ერთობლივად წააგავს ადამიანის მარჯვენა ხელს. დნმ აკავშირებს სამი დომენის მიერ წარმოქმნილ პატარა ჩაღრმავებაში. კატალიზური ცენტრის საფუძველი იქმნება კონსერვატიული ამინომჟავების მოტივებით პალმის დომენში. "თითები" სწორად განათავსებენ მატრიცას აქტიურ ცენტრში. „ცერი“ აკავშირებს დნმ-ს ფერმენტის გასასვლელში და იწვევს მაღალ პროცესურობას. აქტიურ ცენტრში სამივე დომენის ყველაზე მნიშვნელოვანი კონსერვირებული რეგიონები გაერთიანებულია და ქმნის უწყვეტ ზედაპირს. ეგზონუკლეაზას აქტივობა განლაგებულია დამოუკიდებელ დომენში თავისი კატალიზური ადგილით. N-ტერმინალური დომენი ჩასმულია ეგზონუკლეაზას დომენში.
დნმ პოლიმერაზა I-ის მახასიათებლები.
მონაწილეობს დაზიანებული დნმ-ის შეკეთებაში, ასევე თამაშობს დამხმარე როლს დნმ-ის რეპლიკაციაში - აგრძელებს თარგის ჯაჭვთან დაწყვილებულ ჯაჭვის 3'-ბოლოს და საშუალებას გაძლევთ შეავსოთ ხარვეზები m/d ჩამორჩენილი ძაფების ფრაგმენტებით, აგრძელებს ოკაზაკის ფრაგმენტები 3'-ბოლოებიდან, ერთდროულად ამოღებულია რნმ-რიბონუკლეოზიდების თესლები, კატასთან ერთად. ოკაზაკის თითოეული სეგმენტი იწყება. დნმ პოლიმერაზა I-ს შეუძლია გააფართოვოს ერთი ჯაჭვის 3' ბოლო ორჯაჭვიანი დნმ-ის შესვენების დროს და ამოიღოს ნუკლეოტიდები იმავე შესვენების 5' ბოლოდან (ნიკის ტრანსლაცია), რომელიც მნიშვნელოვან როლს ასრულებს აღდგენის სისტემაში.
დნმ პოლიმერი I დომინირებს ყველა დანარჩენში. ეს არის დაახლოებით 103 kDa პოლიპეპტიდი, რომელიც შეიძლება დაიყოს 2 ნაწილად: C-ტერმინალური ფრაგმენტი, 68 kDa, Klenow ფრაგმენტი, აქვს პოლიმერაზული და 3'->5" ეგზონუკლეაზას აქტივობა; N-ტერმინალური ფრაგმენტი, 35 kDa, აქვს. 5'- >3" ეგზონუკლეაზას აქტივობა.
ჰოლოენზიმი, დნმ პოლიმერაზა III, რეპლიზომი.
ჰოლოენზიმი არის კომპლექსი 1 900 კდალ მასით, რომელიც შეიცავს 10 ცილას დაყოფილია 4 ტიპის ქვეკომპლექსად:
α, ξ, θ. შეიცავს კატალიზური ბირთვის 2 ასლს. α - დნმ პოლიმერაზას აქტივობა, ξ - 3'-ეგზონუკლეაზას აქტივობა, θ - ასტიმულირებს ეგზონუკლეაზას.
შეიცავს 2 ქვეერთეულს τ (tau) - ემსახურება მინიმალური ფერმენტის კატალიზური აქტივობით (α) დაკავშირებას.
დამჭერის (სამაგრის) 2 ეგზემპლარი - პასუხისმგებელია დნმ-ის შაბლონებზე მინიმალური ფერმენტის შენარჩუნებაზე. თითოეული შედგება β ქვეერთეულების ჰომოდიმერისგან. მთავარი როლი არის ფერმენტის მატრიციდან გამოყოფის შანსის მინიმუმამდე შემცირება კოპირების პროცესის დასრულებამდე.
γ არის 5 ცილის ჯგუფი, კატა. ფორმირება clamp-loader - მოწყობილობა დნმ-ის მატრიცაზე დამჭერის დასაყენებლად. შედგება 2 δ, 1 γ, 1 ψ და 1 χ - ქვედანაყოფი.

რეპლისომა - მრავალფერმენტული კომპლექსი ბაქტერიების რეპლიკაციის ჩანგალში, რომელიც ახორციელებს ნახევრად კონსერვატიული რეპლიკაციის პროცესს; შეიცავს დნმ პოლიმერაზას და უამრავ სხვა ცილას.
ეუკარიოტული დნმ პოლიმერაზები.
დნმ პოლიმერაზა α - იწყებს ახალი და ჩამორჩენილი ჯაჭვის სინთეზს. ასოცირდება β-ქვეგანყოფილებასთან და ორ პატარა ცილასთან პრიმაზის აქტივობით, ამიტომ მას შეუძლია ჯაჭვების ხელახალი სინთეზირება. 2 ფუნქცია: პრაიმინგი და დრეკადობა = α-პრიმაზა.
დნმ პოლიმერაზა δ - აგრძელებს წამყვან ჯაჭვს
დნმ პოლიმერაზა ξ - მონაწილეობს ჩამორჩენილი ჯაჭვის სინთეზში
დნმ ლიგაზები.
აუცილებელია დნმ-ის ჯაჭვების დასაკავშირებლად რეპლიკაციის, შეკეთების, რეკომბინაციის დროს. E. coli და T4 ფაგის დნმ ლიგაზები არის ერთი პეპტიდი, რომელსაც შეუძლია დააკავშიროს ორი განსხვავებული დუპლექსის ფრაგმენტის ბოლოები ან ხაზოვანი ან წრიული დნმ-ის ჯაჭვების გატეხილი ბოლოები. ამრიგად, დნმ-ის ხაზოვანი და წრიული დუპლექსური მოლეკულები შეიძლება ჩამოყალიბდეს დნმ ლიგაზების დახმარებით.
დნმ ჰელიკაზები.
ახორციელებს ჯაჭვების გაშლას ატფ ჰიდროლიზის ენერგიის გამოყენებით. ის ფუნქციონირებს, როგორც კომპლექსის ნაწილი, რომელიც მოძრაობს რეპლიკაციის ჩანგალს და იმეორებს გადაუხვევ ჯაჭვებს. რამდენიმე ზელიკასს შეუძლია ერთად იმუშაოს სიჩქარის გასაზრდელად.
SSB- ცილები.
ერთჯაჭვიანი შემაკავშირებელი ცილები ახდენენ სპირალის დესტაბილიზაციას, აკავშირებენ ერთჯაჭვიან რეგიონს, რითაც ასტაბილურებენ მას, ე.ი. დააფიქსირეთ ერთჯაჭვიანი დნმ-ის მონაკვეთი.
დნმ ტოპოიზომერაზებიმედაII, გირაზ.
როდესაც დნმ არ გადაუხვევია, მოლეკულა ბრუნავს - ცვლილება მეორად და მესამეულ სტრუქტურებში. ეს პროცესები კატალიზებულია ფერმენტების ჯგუფის მიერ, რომელსაც ეწოდება ტოპოიზომერაზები. ისინი ახდენენ დნმ-ში ერთ და ორჯაჭვიან რღვევებს, რაც საშუალებას აძლევს ნუკლეინის მჟავის მოლეკულას ბრუნოს და გახდეს მატრიცა. მოქმედების მექანიზმის მიხედვით გამოყოფენ პირველი (I) და მეორე (II) ტიპის ტოპოიზომერაზებს.
I ტიპის ტოპოიზომერაზები (E. coli-ში - swivelase) - შეჰყავთ ერთჯაჭვიანი რღვევა დნმ-ის მოლეკულაში, II ტიპის ტოპოიზომერაზები (E. coli-ში - გირაზა) - ახორციელებენ ორჯაჭვიანი დნმ-ის გაწყვეტას და დნმ-ის ჯაჭვების გადატანას დნმ-ის მოლეკულაში. უფსკრული, რასაც მოჰყვება მათი ჯვარედინი კავშირი. ამავდროულად, მათი ფუნქციების შესრულებისას, ტოპიზომერაზები რჩება ასოცირებული დნმ-ის მოლეკულასთან. ამ პროცესებში ტოპოიზომერაზები იყენებენ ტიროზინის ნარჩენს, რომელიც ახორციელებს ნუკლეოფილურ შეტევას დნმ-ის ფოსფატის ჯგუფზე ფოსფოტიროზინის წარმოქმნით. შედეგად, ფერმენტები კოვალენტურად უკავშირდება დნმ-ის 5' ან 3' ბოლოებს შესვენების ადგილზე. ასეთი კოვალენტური ბმის წარმოქმნა გამორიცხავს ენერგიის დახარჯვის აუცილებლობას ფოსფოდიესტერული ბმის აღდგენისას რეაქციის ბოლო ეტაპებზე ერთჯაჭვიანი რღვევის დროს. I ტიპის დნმ ტოპოიზომერაზებს აქვთ ერთი კატალიზური ტიროზინის ნარჩენი თითო მონომერული ცილის მოლეკულაზე, ხოლო დნმ ტოპოიზომერაზა II დიმერები შეიცავს ერთ კატალიზურ ნარჩენს თითო ქვეერთეულზე, რაც ქმნის დნმ-ის მოლეკულაში ორჯაჭვიან რღვევას.
ტოპოიზომერაზები მოქმედებენ როგორც საკინძები, მაგრამ მათი მოქმედებები საპირისპიროა. ტოპოიზომერაზები I, რომლებიც არღვევენ წრიული ზეხვეული დნმ-ის ერთ-ერთ ჯაჭვს, ხსნიან ჯაჭვებს და ამცირებენ სუპერხვეულების რაოდენობას. ტოპოიზომერაზები II გარდაქმნის მოდუნებულ არაზედახვეულ დახურულ წრიულ დნმ-ს სუპერკოჭად.
რეპლიკაციის ეტაპები: დაწყება, გახანგრძლივება, შეწყვეტა. დნმ-ის ახალი ჯაჭვების ფორმირების დაწყება. პრიმაზი. პრიმოსომა. პროკარიოტებში დნმ-ის რეპლიკაციის შეწყვეტა და ქალიშვილი სპირალის დივერგენცია.
რეპლიკაციის შეწყვეტა ხაზოვან გენომებში. ხაზოვანი ღია დნმ-ის ფრაგმენტის რეპლიკაციის პრობლემა. ტელომერები და ტელომერული გამეორებები, ტელომერული მარყუჟი. ტელომერაზა. ტელომერაზას მექანიზმი. ევკარიოტული დნმ-ის რეპლიკაციის მახასიათებლები. ევკარიოტული რეპლიკონები.
როგორც სხვა უჯრედის მაკრომოლეკულების ბიოსინთეზის შემთხვევაში, რეპლიკაციის პროცესი პირობითად იყოფა: სამი ძირითადი ეტაპი: დაწყება, გახანგრძლივება და შეწყვეტა.
პროკარიოტული რეპლიკაცია
ინიცირება
ყველაზე ხშირად წარმოდგენილია პროკარიოტული ქრომოსომა ერთი ზეგადახვეული წრიული მოლეკულა რეპლიკაციის ერთი ან ორი საწყისით.იმისათვის, რომ დნმ-ის ორი ჯაჭვი გახდეს შაბლონი, ახალი ჯაჭვის სინთეზისთვის აუცილებელია დნმ-ის ჯაჭვები გასწორდეს და დაშორდეს ერთმანეთს. დადგენილია, რომ დნმ-ის ჯაჭვები იხსნება არა მთელ სიგრძეზე, არამედ მოკლე მონაკვეთზე. აქ იქმნება რეპლიკაციის ჩანგალი - დნმ-ის დუბლირების ადგილი.
როდესაც დნმ არ გადაუხვევია, მოლეკულა ბრუნავს - ცვლილება მეორად და მესამეულ სტრუქტურებში. ეს პროცესები კატალიზებულია ფერმენტების ჯგუფის მიერ ე.წ ტოპოიზომერაზები . ისინი ახდენენ დნმ-ში ერთ და ორჯაჭვიან რღვევებს, რაც საშუალებას აძლევს ნუკლეინის მჟავის მოლეკულას ბრუნოს და გახდეს მატრიცა. მოქმედების მექანიზმის მიხედვით გამოყოფენ პირველი (I) და მეორე (II) ტიპის ტოპოიზომერაზებს.
მშობელი დნმ-ის მოლეკულის დაუხვეველ მონაკვეთზე, საიდანაც იწყება რეპლიკაცია და რომელსაც რეპლიკაციის საწყისს (ანუ წარმოშობას, oriC) უწოდებენ, ინიციატორი ცილები „დაჯდებიან“. oriC-ში რეპლიკაციის დაწყება იწყება კომპლექსის წარმოქმნით, რომელიც მოიცავს ექვს პროტეინს DnaA, DnaB, DnaC, HU, გირაზას და SSB.
ცილები პირველ რიგში უკავშირდებიან ცხრა ნუკლეოტიდულ თანმიმდევრობას DnaA , რომლებიც ქმნიან დიდ აგრეგატს. წარმოშობის დნმ გარშემორტყმულია მას და დნმ-ის ძაფები გამოყოფილია სამი 13-მერიანი თანმიმდევრობის რეგიონში. შემდეგ ეტაპზე, DnaB (ჰელიკაზა) და DnaC ცილები მიმაგრებულია, ქმნიან 480 kDa აგრეგატს 6 ნმ რადიუსით. ჰელიკაზა/ DnaB უზრუნველყოფს წყალბადის ბმების გაწყვეტას აზოტოვან ფუძეებს შორის დნმ-ის ორმაგ ჯაჭვში, რაც იწვევს მის დენატურაციას, ე.ი. ძაფების განსხვავება.
დნმ-ის ორმაგი სპირალის გასწორებისა და დენატურაციის შედეგად წარმოიქმნება Y- ფორმის რეპლიკაციის ჩანგალი (ნახ. სწორედ ამ რეპლიკაციის ჩანგლით დნმ-პოლიმერაზები ასინთეზირებენ ქალიშვილის დნმ-ის მოლეკულებს. დნმ-ის ასეთი ნაჭერი ჰგავს ბუშტს ან „თვალს“ გაუმეორებელ დნმ-ში. რეპლიკაციის "თვალები" იქმნება იმ ადგილებში, სადაც განლაგებულია რეპლიკაციის წარმოშობის წერტილები. როდესაც დნმ-ის ჯაჭვები გათიშულია, მოლეკულა საკმაოდ მოძრავი ხდება. მოქმედების გამო გამორიცხულია ყველა შესაძლო დარღვევა ერთჯერადი სქემების სტრუქტურაში SSB ცილები
(ერთჯაჭვიანი დნმ-ის დამაკავშირებელი პროტეინები ან სპირალის დესტაბილიზაციის პროტეინები), რომლებიც დნმ-ის ცალკეულ ჯაჭვებთან შეკავშირებით ხელს უშლიან მათ ერთმანეთთან შეკვრას. 
არის მემკვიდრეობითი მოლეკულა, მაშინ იმისათვის, რომ გააცნობიეროს ეს თვისება, მან ზუსტად უნდა დააკოპიროს საკუთარი თავი და ამით შეინახოს ორიგინალური დნმ-ის მოლეკულაში არსებული ყველა ინფორმაცია ნუკლეოტიდების კონკრეტული თანმიმდევრობის სახით. ამას უზრუნველყოფს სპეციალური პროცესი, რომელიც წინ უსწრებს სხეულის რომელიმე უჯრედის გაყოფას, რომელსაც დნმ-ის რეპლიკაცია ეწოდება.
დნმ-ის რეპლიკაციის არსი იმაში მდგომარეობს, რომ სპეციალური ფერმენტი არღვევს სუსტ წყალბადურ ბმებს, რომლებიც აკავშირებენ ორი ჯაჭვის ნუკლეოტიდებს. შედეგად, დნმ-ის ჯაჭვები გათიშულია და თავისუფალი აზოტოვანი ფუძეები „გამოდის“ თითოეული ჯაჭვიდან (ე.წ. რეპლიკაციის ჩანგლის გამოჩენა). სპეციალური ფერმენტი, დნმ პოლიმერაზა, იწყებს მოძრაობას დნმ-ის თავისუფალი ჯაჭვის გასწვრივ მე-5-დან მე-3-ბოლომდე (წამყვანი ჯაჭვი), რაც ეხმარება უჯრედში მუდმივად სინთეზირებულ თავისუფალ ნუკლეოტიდების შეერთებას ახლად სინთეზირებულის 3" ბოლოში. დნმ-ის ჯაჭვი დნმ-ის მეორე ჯაჭვზე (ჩამორჩენილი ჯაჭვი) წარმოიქმნება ახალი დნმ მცირე სეგმენტების სახით, რომელიც შედგება 1000-2000 ნუკლეოტიდისგან (ოკაზაკის ფრაგმენტები).
ამ ჯაჭვის დნმ-ის ფრაგმენტების რეპლიკაციის დასაწყებად საჭიროა რნმ-ის მოკლე ფრაგმენტების სინთეზი (რნმ-ის დამახასიათებელი ნიშნები ქვემოთ იქნება განხილული) თესლის სახით, რისთვისაც გამოიყენება სპეციალური ფერმენტი - რნმ პოლიმერაზა (პრიმაზა). შემდგომში, რნმ პრაიმერები ამოღებულია და დნმ შეჰყავთ ხარვეზებში დნმ პოლიმერაზა I-ის გამოყენებით. ამრიგად, დნმ-ის თითოეული ჯაჭვი გამოიყენება როგორც შაბლონი ან შაბლონი დამატებითი ჯაჭვის შესაქმნელად, ხოლო დნმ-ის რეპლიკაცია ნახევრად კონსერვატიულია (ანუ ერთი ჯაჭვი. ახალ დნმ-ში მოლეკულა არის „ძველი“, მეორე კი ახალი).
უჯრედში წამყვანი და ჩამორჩენილი ძაფების გასამრავლებლად გამოიყენება სხვადასხვა ფერმენტები. რეპლიკაციის შედეგად წარმოიქმნება ორი ახალი აბსოლუტურად იდენტური დნმ-ის მოლეკულა, რომლებიც ასევე იდენტურია ორიგინალური დნმ-ის მოლეკულისა მისი რედუპლიკაციის დაწყებამდე (დნმ-ის რეპლიკაციის პროცესი უფრო დეტალურად არის ნაჩვენები ნახ. 3.5). დნმ პოლიმერაზა, ისევე როგორც ნებისმიერი სხვა ფერმენტი, მნიშვნელოვნად აჩქარებს დამატებითი ნუკლეოტიდების დამატების პროცესს თავისუფალ დნმ-ის ჯაჭვში, თუმცა, ადენინის ქიმიური მიდრეკილება თიმინთან და ციტოზინის მიმართ გუანინთან იმდენად მაღალია, რომ ისინი ერწყმის ერთმანეთს. დნმ პოლიმერაზას არარსებობა მარტივი რეაქციის ნარევებში.
შეიძლება ითქვას, გარკვეულწილად გამარტივებით, რომ დნმ-ის მოლეკულის ზუსტი გაორმაგების ფენომენი, რომელიც ემყარება ამ მოლეკულის ფუძეების კომპლემენტარულობას, არის მემკვიდრეობის მოლეკულური საფუძველი. ადამიანებში დნმ-ის რეპლიკაციის სიჩქარე შედარებით ნელია და კვირები დასჭირდება ადამიანის ნებისმიერი ქრომოსომის დნმ-ის რეპლიკაციას, თუ რეპლიკაცია დაიწყება ერთი წერტილიდან. სინამდვილეში, ნებისმიერი ქრომოსომის დნმ-ის მოლეკულაში და თითოეული ადამიანის ქრომოსომა შეიცავს მხოლოდ ერთ დნმ-ის მოლეკულას, არის რეპლიკაციის დაწყების მრავალი ადგილი (რეპლიკონი). თითოეული რეპლიკონიდან რეპლიკაცია მიმდინარეობს ორივე მიმართულებით, სანამ მეზობელი რეპლიკონები არ გაერთიანდებიან. ამიტომ, თითოეულ ქრომოსომაში დნმ-ის რეპლიკაცია შედარებით სწრაფად მიმდინარეობს.
რეპლიკაცია (ლათ. replicatio - განახლება) - ქალიშვილის დნმ-ის მოლეკულის სინთეზის პროცესი მშობელი დნმ-ის მოლეკულის მატრიცაზე. დედა უჯრედის შემდგომი გაყოფის დროს, თითოეული შვილობილი უჯრედი იღებს დნმ-ის მოლეკულის ერთ ასლს, რომელიც იდენტურია ორიგინალური დედა უჯრედის დნმ-ის. ეს პროცესი უზრუნველყოფს გენეტიკური ინფორმაციის ზუსტ გადაცემას თაობიდან თაობას. დნმ-ის რეპლიკაცია ხორციელდება რთული ფერმენტული კომპლექსით, რომელიც შედგება 15-20 განსხვავებული ცილისგან, რომელსაც ეწოდება რეპლიზომი (ინგლისური რეპლიზომი).
დნმ-ის რეპლიკაცია არის საკვანძო მოვლენა უჯრედების გაყოფის პროცესში. მნიშვნელოვანია, რომ გაყოფის მომენტისთვის დნმ მთლიანად განმეორდება და მხოლოდ ერთხელ. ამას უზრუნველყოფს დნმ-ის რეპლიკაციის რეგულირების გარკვეული მექანიზმები.
რეპლიკაცია ხდება სამი ეტაპი:
1. რეპლიკაციის დაწყება
2. დრეკადობა
3. რეპლიკაციის შეწყვეტა.
რეპლიკაცია რეგულირდება ძირითადად დაწყების ეტაპზე. ამის განხორციელება საკმაოდ მარტივია, რადგან რეპლიკაცია შეიძლება დაიწყოს არა რომელიმე დნმ-ის სეგმენტიდან, არამედ მკაცრად განსაზღვრულიდან, რომელსაც ეწოდება რეპლიკაციის დაწყების ადგილი. გენომში შეიძლება იყოს მხოლოდ ერთი ან ბევრი ასეთი ადგილი. რეპლიკაციის დაწყების ადგილის ცნება მჭიდროდ არის დაკავშირებული რეპლიკონის ცნებასთან. რეპლიკონი არის დნმ-ის მონაკვეთი, რომელიც შეიცავს რეპლიკაციის დაწყების ადგილს და მრავლდება ამ ადგილიდან დნმ-ის სინთეზის დაწყების შემდეგ. ბაქტერიული გენომები, როგორც წესი, არის ერთი რეპლიკონი, რაც ნიშნავს, რომ მთელი გენომის რეპლიკაცია არის რეპლიკაციის დაწყების მხოლოდ ერთი აქტის შედეგი.
ევკარიოტული გენომები (ისევე, როგორც მათი ცალკეული ქრომოსომა) შედგება დიდი რაოდენობით დამოუკიდებელი რეპლიკებისგან, რაც მნიშვნელოვნად ამცირებს ცალკეული ქრომოსომის რეპლიკაციის მთლიან დროს. მოლეკულურ მექანიზმებს, რომლებიც აკონტროლებენ რეპლიკაციის დაწყების რაოდენობას თითოეულ ადგილზე უჯრედის გაყოფის ციკლზე, ეწოდება ასლის რაოდენობის კონტროლი. ქრომოსომული დნმ-ის გარდა, ბაქტერიული უჯრედები ხშირად შეიცავს პლაზმიდებს, რომლებიც ინდივიდუალური რეპლიკონებია. პლაზმიდებს აქვთ ასლების რაოდენობის კონტროლის საკუთარი მექანიზმები: მათ შეუძლიათ უზრუნველყონ პლაზმიდის მხოლოდ ერთი ასლის სინთეზი უჯრედულ ციკლზე, ან ათასობით ასლი.
რეპლიკაცია იწყება რეპლიკაციის დაწყების ადგილზე, დნმ-ის ორმაგი სპირალის გახსნით, რეპლიკაციის ჩანგლის წარმოქმნით, დნმ-ის პირდაპირი რეპლიკაციის ადგილზე. თითოეულ საიტს შეუძლია შექმნას ერთი ან ორი რეპლიკაციის ჩანგალი, იმისდა მიხედვით, რეპლიკაცია არის ცალმხრივი თუ ორმხრივი. ორმხრივი რეპლიკაცია უფრო ხშირია. რეპლიკაციის დაწყებიდან გარკვეული პერიოდის შემდეგ, ელექტრონული მიკროსკოპი შეიძლება გამოყენებულ იქნას რეპლიკაციის თვალის დასაკვირვებლად - ქრომოსომის რეგიონი, სადაც დნმ უკვე გამრავლებულია, გარშემორტყმული არარეპლიკაციური დნმ-ის უფრო გაფართოებული უბნებით.
რეპლიკაციის ჩანგალში დნმ აკოპირებს დიდ ცილის კომპლექსს (რეპლიზომას), რომლის ძირითადი ფერმენტი არის დნმ პოლიმერაზა. რეპლიკაციის ჩანგალი მოძრაობს დაახლოებით 100000 ბაზის წყვილი წუთში პროკარიოტებში და 500-5000 ევკარიოტებში.
რეპლიკაციის მოლეკულური მექანიზმი:
ფერმენტები (ჰელიკაზა, ტოპოიზომერაზა) და დნმ-ის დამაკავშირებელი ცილები ხსნიან დნმ-ს, ატარებენ მატრიქსს განზავებულ მდგომარეობაში და ატრიალებენ დნმ-ის მოლეკულას. რეპლიკაციის სისწორეს უზრუნველყოფს დამატებითი ბაზის წყვილების ზუსტი შეხამება და დნმ პოლიმერაზას აქტივობა, რომელსაც შეუძლია შეცდომის ამოცნობა და გამოსწორება. ევკარიოტებში რეპლიკაცია ხორციელდება რამდენიმე განსხვავებული დნმ პოლიმერაზას მიერ (პროკარიოტებში დნმ-ის რეპლიკაციისგან განსხვავებით).
დნმ პოლიმერაზა I მოქმედებს ჩამორჩენილ ჯაჭვზე რნმ პრაიმერების მოსაშორებლად და გასუფთავებული დნმ-ის ადგილების წინასწარ რეპლიკაციისთვის. დნმ პოლიმერაზა III არის დნმ-ის რეპლიკაციის მთავარი ფერმენტი, რომელიც ასინთეზებს დნმ-ის წამყვან ჯაჭვს და ოკაზაკის ფრაგმენტებს ჩამორჩენილი ჯაჭვის სინთეზის დროს (ოკაზაკის ფრაგმენტები შედარებით მოკლე დნმ-ის ფრაგმენტებია, რომლებიც წარმოიქმნება ჩამორჩენილ ჯაჭვზე დნმ-ის რეპლიკაციის დროს). შემდეგი, სინთეზირებული მოლეკულები იგრიხება დნმ-ის ზეგადახვევისა და შემდგომი დატკეპნის პრინციპის მიხედვით. სინთეზი ენერგო ინტენსიურია.
დნმ-ის მოლეკულის ჯაჭვები განსხვავდებიან, ქმნიან რეპლიკაციის ჩანგალს და თითოეული მათგანი ხდება შაბლონი, რომელზედაც სინთეზირებულია ახალი დამატებითი ჯაჭვი. შედეგად, წარმოიქმნება ორი ახალი ორჯაჭვიანი დნმ-ის მოლეკულა, რომლებიც იდენტურია მშობელი მოლეკულისა.
რეპლიკაცია- ინფორმაციის გადაცემა დნმ-დან დნმ-ზე, დნმ-ის თვითგამრავლება (დნმ ბიოსინთეზი).
დნმ-ის მოლეკულა შედგება ორი ჯაჭვისგან ორმაგდებაუჯრედების გაყოფის დროს. დნმ-ის გაორმაგებაემყარება იმ ფაქტს, რომ ძაფების ამოღებისას თითოეული ძაფის დასრულება შესაძლებელია დამატებითი ასლი, რითაც მიიღება დნმ-ის მოლეკულის ორი ჯაჭვი, ორიგინალის კოპირება.
რეპლიკაციისთვის საჭირო პირობები: 1.) მატრიცა- დნმ-ის ჯაჭვები. ძაფის გაყოფა ეწოდება რეპლიკაციის ჩანგალი. ის შეიძლება ჩამოყალიბდეს დნმ-ის მოლეკულაში. ისინი მოძრაობენ სხვადასხვა მიმართულებით, ქმნიან რეპლიკაციური თვალი. ევკარიოტების დნმ-ის მოლეკულაში რამდენიმე ასეთი თვალია, თითოეულს ორი ჩანგალი აქვს. 2.) სუბსტრატი. პლასტიკური მასალა არის დეოქსინუკლეოტიდის ტრიფოსფატები: dATP, dGTP, dCTP, dTTP. შემდეგ ისინი იშლება დეოქსინუკლეოტიდური მონოფოსფატები, არაორგანული ფოსფატის ორი მოლეკულა ენერგიის გამოყოფით, ე.ი. ისინი ორივე წყაროა და ენერგია, და პლასტიკური მასალა. 3.) იონები მაგნიუმი. 4.) რეპლიკაციური ფერმენტის კომპლექსი. ა) დნმ ხსნის ცილებს: - დნმ-ა(იწვევს ძაფების დივერგენციას); - ჰელიკაზები(დნმ-ის ჯაჭვის გაწყვეტა) - ტოპოიზომერაზები 1და 2 (განტვირთვა სპირალზე). გაანადგურე (3",5")-ფოსფოდიესტერული ბმები. პროკარიოტებში ტოპოიზომერაზა 2 ე.წ გირაზა. ბ) ცილები, რომლებიც ხელს უშლიან დნმ-ის ჯაჭვების შეერთებას ( SSB ცილები). V) დნმ პოლიმერაზა(კატალიზებს ფოსფოდიესტერული ბმების წარმოქმნას). დნმ პოლიმერაზა მხოლოდ ახანგრძლივებს უკვე არსებულ ჯაჭვს, მაგრამ არ შეუძლია დააკავშიროს ორი თავისუფალი ნუკლეოტიდი. გ) პრიმაზა(აკატალიზებს სინთეზისთვის „თესლის“ წარმოქმნას). ეს არის თავის სტრუქტურაში რნმ პოლიმერაზა, რომელიც აკავშირებს ერთ ნუკლეოტიდებს. ე) დნმ ლიგაზა. 5.) პრაიმერები- "თესლი" გამრავლებისთვის. ეს არის მოკლე ფრაგმენტი რიბონუკლეოტიდის ტრიფოსფატები(2 - 10). პრაიმერის ფორმირება კატალიზებულია პრიმაზა.
რეპლიკაციის ნაბიჯები: 1.) ინიცირება(რეპლიკაციის ჩანგლის ფორმირება); 2.) დრეკადობა(ახალი ძაფების სინთეზი); 3.) პრაიმერის გამორიცხვა; 4.) შეწყვეტა(ორი ბავშვის ჯაჭვის სინთეზის დასრულება).
რეპლიკაციის დაწყება:- არეგულირებს სასიგნალო ცილის მოლეკულებს - ზრდის ფაქტორები;- უზრუნველყოფა ფერმენტებიდა სპეციალური ცილები.
საჭირო ფერმენტები: დნმ ტოპოიზომერაზებიფერმენტები, რომლებიც ხსნიან დნმ-ის სუპერკოპირებს. დნმ ჰელიკაზა- არღვევს წყალბადურ კავშირებს ორჯაჭვიან დნმ-ის მოლეკულაში. შედეგად, ა რეპლიკაციის ჩანგალი (რეპლიკაციური თვალი).

ცილები, რომლებიც უკავშირდებიან დნმ-ის ერთჯაჭვიან ძაფებს, უკავშირდებიან ერთჯაჭვიან დნმ-ს და ხელს უშლიან მათ დამატებით შეერთებას.
რეპლიკაციის გახანგრძლივება.სინთეზის სუბსტრატებია დეოქსინუკლეოზიდის ტრიფოსფატები, მოქმედებს როგორც სამშენებლო მასალა და ენერგიის წყარო.
საჭირო ფერმენტები: დნმ პრიმაზა, რომელიც კატალიზებს რნმ-ის პრაიმერების მოკლე მოლეკულების სინთეზს დნმ პოლიმერაზასთვის. დნმ პოლიმერაზაუზრუნველყოფს ნუკლეოტიდების მზარდ "ახალ" ჯაჭვში ჩართვას, რომელიც ავსებს "ძველს", ანუ შაბლონის ჯაჭვს.
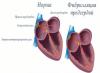
დნმ-ის ახალი ჯაჭვების სინთეზი შეიძლება მხოლოდ მიმართულებით მიმდინარეობდეს 5' ბოლოდან 3' ბოლომდე. ერთ ჯაჭვზე დნმ განუწყვეტლივ სინთეზირდება "წამყვანი" ჯაჭვი, ხოლო მეორეზე ყალიბდება მოკლე ფრაგმენტები - "ჩამორჩენილი" ჯაჭვი (ოკაზაკის ფრაგმენტები).
პრაიმერების მოხსნის შემდეგ დნმ ლიგაზააკერებს ოკაზაკის მოკლე ფრაგმენტებს ( შეწყვეტა).
ინფორმაცია გადაცემულია მატრიცული გზა. ნახევრად კონსერვატიულიდნმ-ის რეპლიკაციის მექანიზმი.
| ჩამორჩენილი ძაფების სინთეზი |
| 3’ |
| 3’ |
| 5’ |
| 5’ |


დნმ არის გენეტიკური ინფორმაციის საიმედო საცავი. მაგრამ ის არა მხოლოდ უსაფრთხოდ უნდა იყოს დაცული, არამედ შთამომავლობასაც გადაეცეს. სახეობების გადარჩენა დამოკიდებულია ამაზე. მშობლებმა ხომ უნდა გადასცენ შვილებს ყველაფერი, რაც მათ ევოლუციის პროცესში მიაღწიეს. მასში ყველაფერია ჩაწერილი: კიდურების რაოდენობით დაწყებული თვალების ფერებამდე. რა თქმა უნდა, მიკროორგანიზმებს ეს ინფორმაცია გაცილებით ნაკლები აქვთ, მაგრამ ის ასევე უნდა იყოს გადაცემული. ამისათვის უჯრედი იყოფა. იმისათვის, რომ გენეტიკური ინფორმაცია ორივე ქალიშვილ უჯრედში მივიდეს, ის უნდა გაორმაგდეს, ამ პროცესს „დნმ-ის რეპლიკაცია“ ეწოდება. ეს ხდება უჯრედის გაყოფამდე, არ აქვს მნიშვნელობა რომელი. ეს შეიძლება იყოს ბაქტერია, რომელმაც გამრავლება გადაწყვიტა. ან ეს შეიძლება იყოს კანის ახალი ზრდა ჭრილობის ადგილზე. დეზოქსირიბონუკლეინის მჟავის დუბლირების პროცესი მკაფიოდ უნდა დარეგულირდეს და დასრულდეს უჯრედების გაყოფის დაწყებამდე.
სად ხდება გაორმაგება?
დნმ-ის რეპლიკაცია ხდება უშუალოდ ბირთვში (ევკარიოტებში) ან ციტოპლაზმაში (პროკარიოტებში). ნუკლეინის მჟავა შედგება ნუკლეოტიდებისგან - ადენინი, თიმინი, ციტოზინი და გუანინი. მოლეკულის ორივე ჯაჭვი აგებულია კომპლემენტარობის პრინციპით: ერთ ჯაჭვში ადენინი შეესაბამება თიმინს, ხოლო გუანინი ციტოზინს. მოლეკულის გაორმაგება ისე უნდა მოხდეს, რომ ასული ხვეულებში შენარჩუნდეს კომპლემენტარობის პრინციპი.

რეპლიკაციის დაწყება – დაწყება
დეოქსირიბონუკლეინის მჟავა არის ორჯაჭვიანი სპირალი. დნმ-ის რეპლიკაცია ხდება ბავშვის ძაფების აგების დასრულებით თითოეული მშობელი ჯაჭვის გასწვრივ. იმისათვის, რომ ეს სინთეზი გახდეს შესაძლებელი, სპირალები უნდა „გაიშალოს“ და ჯაჭვები განცალკევდეს ერთმანეთისგან. ამ როლს ასრულებს ჰელიკაზა - ის ხსნის დეზოქსირიბონუკლეინის მჟავას სპირალს, ბრუნავს დიდი სიჩქარით. დნმ-ის დუბლირების დასაწყისი ვერცერთი ადგილიდან ვერ დაიწყება, ასეთი რთული პროცესი მოითხოვს მოლეკულის გარკვეულ ნაწილს – რეპლიკაციის დაწყების ადგილს. მას შემდეგ, რაც განისაზღვრა დუბლირების საწყისი წერტილი და ჰელიკაზა იწყებს მუშაობას სპირალის გასახსნელად, დნმ-ის ძაფები გვერდებზე გადადის და ქმნის რეპლიკაციის ჩანგალს. მათზე დნმ პოლიმერაზები ზის. სწორედ ისინი მოახდენენ ბავშვის ჯაჭვების სინთეზს.

დრეკადობა

დეზოქსირიბონუკლეინის მჟავის ერთ მოლეკულაში შეიძლება ჩამოყალიბდეს 5-დან 50-მდე რეპლიკაციის ჩანგალი. ქალიშვილი ჯაჭვების სინთეზი ერთდროულად ხდება მოლეკულის რამდენიმე რეგიონში. მაგრამ დამატებითი ნუკლეოტიდების აგების დასრულება ადვილი არ არის. ნუკლეინის მჟავას ჯაჭვები ერთმანეთის ანტიპარალელურია. მშობელი ჯაჭვების განსხვავებული ორიენტაცია გავლენას ახდენს გაორმაგებაზე, რამაც გამოიწვია დნმ-ის რეპლიკაციის რთული მექანიზმი. ერთ-ერთ ჯაჭვს ბავშვის ჯაჭვი უწყვეტად ავსებს და ლიდერს უწოდებენ. ეს სწორია, რადგან პოლიმერაზასთვის ძალიან მოსახერხებელია თავისუფალი ნუკლეოტიდის მიმაგრება წინა 3'-OH ბოლოზე. ასეთი სინთეზი მიმდინარეობს უწყვეტად, განსხვავებით მეორე ჯაჭვის პროცესისგან.
ჩამორჩენილი ჯაჭვი, ო'კაზაკების ფრაგმენტები
სირთულეები წარმოიქმნება სხვა ჯაჭვთან დაკავშირებით, რადგან არსებობს თავისუფალი 5'-ბოლო, რომელზეც შეუძლებელია თავისუფალი ნუკლეოტიდის მიმაგრება. შემდეგ დნმ პოლიმერაზა მოქმედებს მეორე მხარეს. ბავშვის ძაფების დასასრულებლად, იქმნება პრაიმერი, რომელიც ავსებს მშობელს. იგი იქმნება თავად რეპლიკაციის ჩანგალზე. მცირე ნაწილის სინთეზი იწყება მასთან, მაგრამ უკვე "სწორი" გზის გასწვრივ - ნუკლეოტიდების დამატება ხდება 3'-ბოლომდე. ამრიგად, ჯაჭვის დასრულება მეორე ქალიშვილის სპირალში ხდება შეუწყვეტლად და აქვს მიმართულება რეპლიკაციის ჩანგლის მოძრაობის საწინააღმდეგოდ. ამ ფრაგმენტებს ეწოდა ო'კაზაკის ფრაგმენტები და სიგრძეში დაახლოებით 100 ნუკლეოტიდია. მას შემდეგ, რაც ფრაგმენტი დასრულდება წინა მზა ნაჭერზე, პრაიმერები იჭრება სპეციალური ფერმენტით, ამოჭრილი ადგილი ივსება დაკარგული ნუკლეოტიდებით.
შეწყვეტა
გაორმაგება სრულდება, როდესაც ორივე ჯაჭვი დაასრულებს შვილებს და ო'კაზაკების ყველა ფრაგმენტი ერთად შეიკერება. ევკარიოტებში დნმ-ის რეპლიკაცია მთავრდება, როდესაც რეპლიკაციის ჩანგლები ერთმანეთს ხვდებიან. პროკარიოტებში კი ეს მოლეკულა წრიულია და მისი გაორმაგების პროცესი ჯაჭვის გაწყვეტის გარეშე ხდება. გამოდის, რომ ყველა დეზოქსირიბონუკლეინის მჟავა არის ერთი დიდი რეპლიკონი. და გაორმაგება მთავრდება, როცა რეპლიკაციის ჩანგლები რგოლის მოპირდაპირე მხარეს ხვდება. რეპლიკაციის დასრულების შემდეგ, ძირითადი დეზოქსირიბონუკლეინის მჟავის ორივე ჯაჭვი უნდა იყოს უკუკავშირი, რის შემდეგაც ორივე მოლეკულა გადაუგრიხეს ზებრკოლების წარმოქმნით. შემდეგი, ორივე დნმ-ის მოლეკულის მეთილაცია ხდება ადენინზე -GATC- ადგილზე. ეს არ წყვეტს ჯაჭვებს და არ უშლის ხელს მათ კომპლემენტარულობას. ეს აუცილებელია მოლეკულების ქრომოსომებში დასაკეცი, ასევე გენის წაკითხვის რეგულირებისთვის.
გამეორების სიჩქარე და სიზუსტე

დნმ-ის დუბლირების (დრეკადობის) მეორე ეტაპი ხდება წამში დაახლოებით 700 ნუკლეოტიდის სიჩქარით. თუ გავიხსენებთ, რომ ნუკლეინის მჟავის ერთ შემობრუნებაზე 10 წყვილი მონომერია, მაშინ გამოდის, რომ „გახვევის“ დროს მოლეკულა ბრუნავს წამში 70 ბრუნის სიხშირით. შედარებისთვის: გამაგრილებლის ბრუნვის სიჩქარე კომპიუტერის სისტემურ ერთეულში არის დაახლოებით 500 ბრუნი წამში. მაგრამ მიუხედავად მაღალი მაჩვენებლებისა, დნმ პოლიმერაზა თითქმის არასოდეს არის არასწორი. ყოველივე ამის შემდეგ, ის უბრალოდ ირჩევს დამატებით ნუკლეოტიდებს. მაგრამ შეცდომის დაშვების შემთხვევაშიც კი, დნმ პოლიმერაზა ცნობს მას, დგას ნაბიჯი უკან, ამოიღებს არასწორ მონომერს და ცვლის მას სწორით. დნმ-ის რეპლიკაციის მექანიზმი ძალიან რთულია, მაგრამ ჩვენ შევძელით ძირითადი პუნქტები. მნიშვნელოვანია გვესმოდეს მისი მნიშვნელობა როგორც მიკროორგანიზმებისთვის, ასევე მრავალუჯრედიანი ორგანიზმებისთვის.
დნმ-ის რეპლიკაციაარის მისი გაორმაგების პროცესი უჯრედების გაყოფამდე. ზოგჯერ მას უწოდებენ "დნმ-ის რეპლიკაციას". გაორმაგება ხდება უჯრედული ციკლის ინტერფაზის S ფაზაში.
ცხადია, ველურ ბუნებაში გენეტიკური მასალის თვითკოპირება აუცილებლობაა. მხოლოდ ამ გზით შეიძლება უჯრედების გაყოფის დროს წარმოქმნილი შვილობილი უჯრედები შეიცავდეს იმდენ დნმ-ს, რამდენიც თავდაპირველად იყო თავდაპირველში. რეპლიკაციის წყალობით, სტრუქტურისა და მეტაბოლიზმის ყველა გენეტიკურად დაპროგრამებული მახასიათებელი გადაეცემა რამდენიმე თაობას.
უჯრედის გაყოფის პროცესში, დნმ-ის თითოეული მოლეკულა იდენტური წყვილიდან მიდის თავის ქალიშვილ უჯრედში. ეს უზრუნველყოფს მემკვიდრეობითი ინფორმაციის ზუსტ გადაცემას.
დნმ-ის სინთეზის დროს ენერგია იხარჯება, ანუ ეს არის ენერგიის მომხმარებელი პროცესი.
დნმ-ის რეპლიკაციის მექანიზმი
თავად დნმ-ის მოლეკულა (გაორმაგების გარეშე) არის ორმაგი სპირალი. რედუპლიკაციის პროცესში წყალბადის ბმები მის ორ დამატებით ჯაჭვს შორის იშლება. და თითოეულ ცალკეულ ჯაჭვზე, რომელიც ახლა შაბლონ-მატრიცას ემსახურება, აგებულია ახალი დამატებითი ჯაჭვი. ამრიგად, წარმოიქმნება დნმ-ის ორი მოლეკულა. თითოეული მათგანი იღებს თითო ჯაჭვს დედის დნმ-დან, მეორე კი ახლად სინთეზირებულია. ამიტომ, დნმ-ის რეპლიკაციის მექანიზმი არის ნახევრად კონსერვატიული(ერთი ჯაჭვი ძველი, ერთი ახალი). რეპლიკაციის ეს მექანიზმი დადასტურდა 1958 წელს.
დნმ-ის მოლეკულაში ძაფები ანტიპარალელურია. ეს ნიშნავს, რომ ერთი ღერი მიდის მიმართულებით 5" ბოლოდან 3-მდე", ხოლო დამატებითი ერთი - პირიქით. ნომრები 5 და 3 მიუთითებს ნახშირბადის ატომების რაოდენობას დეზოქსირიბოზაში, რომელიც თითოეული ნუკლეოტიდის ნაწილია. ნუკლეოტიდები დაკავშირებულია ამ ატომების მეშვეობით ფოსფოდიესტერული ბმებით. და სადაც ერთ ჯაჭვს აქვს 3 "შეერთება, მეორეს აქვს 5", რადგან თავდაყირა დგას, ანუ მიდის მეორე მიმართულებით. სიცხადისთვის, შეგიძლიათ წარმოიდგინოთ, რომ მერხთან მჯდომი პირველკლასელივით ხელზე აჭერით ხელს.
მთავარი ფერმენტი, რომელიც ახორციელებს დნმ-ის ახალი ჯაჭვის გაფართოებას, შეუძლია ამის გაკეთება მხოლოდ ერთი მიმართულებით. კერძოდ: ახალი ნუკლეოტიდის მიმაგრება მხოლოდ 3"-ის ბოლოზე. ამრიგად, სინთეზი შეიძლება წავიდეს მხოლოდ 5"-დან 3"-მდე მიმართულებით.
ჯაჭვები ანტიპარალელურია, რაც იმას ნიშნავს, რომ სინთეზი მათკენ უნდა წავიდეს სხვადასხვა მიმართულებით. თუ დნმ-ის ჯაჭვები ჯერ მთლიანად განსხვავდებოდა და შემდეგ მათზე აშენდებოდა ახალი დამატებითი, მაშინ ეს პრობლემა არ იქნებოდა. სინამდვილეში, ჯაჭვები გარკვეულწილად განსხვავდება რეპლიკაციის საწყისი წერტილებიდა მატრიცების ამ ადგილებში დაუყოვნებლივ იწყება სინთეზი.
Ე. წ რეპლიკაციის ჩანგლები. ამავდროულად, ერთ მშობელ ჯაჭვზე, სინთეზი მიმდინარეობს ჩანგლის დივერგენციის მიმართულებით და ეს სინთეზი ხდება განუწყვეტლივ, შესვენების გარეშე. მეორე შაბლონზე სინთეზი მიმდინარეობს ორიგინალური დნმ-ის ჯაჭვების განსხვავების მიმართულებით საპირისპირო მიმართულებით. მაშასადამე, ასეთი საპირისპირო სინთეზი შეიძლება მოხდეს მხოლოდ ნაწილებად, რომლებიც ე.წ ოკაზაკის ფრაგმენტები. მოგვიანებით ასეთი ფრაგმენტები ერთმანეთში „იკერება“.
ბავშვის ჯაჭვი, რომელიც მუდმივად მეორდება, ეწოდება წამყვანი ან წამყვანი. ის, რომელიც სინთეზირებულია ოკაზაკის ფრაგმენტებით - ჩამორჩენილი, ან ჩამორჩენილირადგან ფრაგმენტული რეპლიკაცია უფრო ნელია.
დიაგრამაში, მშობელი დნმ-ის ძაფები თანდათან განსხვავდება იმ მიმართულებით, რომლითაც სინთეზირებულია წამყვანი ქალიშვილის ჯაჭვი. ჩამორჩენილი ჯაჭვის სინთეზი მიდის დივერგენციის საპირისპირო მიმართულებით, ამიტომ ის უნდა შესრულდეს ნაწილებად.
მთავარი დნმ-ის სინთეზის ფერმენტის (პოლიმერაზას) კიდევ ერთი თვისებაა ის, რომ მას არ შეუძლია თავად დაიწყოს სინთეზი, მხოლოდ გააგრძელოს. Მას სჭირდება პრაიმერი ან პრაიმერი. ამიტომ, მცირე დამატებითი რნმ-ის რეგიონი ჯერ სინთეზირდება მშობლის ჯაჭვზე, შემდეგ ჯაჭვი გაფართოვდება პოლიმერაზას გამოყენებით. მოგვიანებით, პრაიმერები ამოღებულია და ხვრელები აშენებულია.
დიაგრამაში თესლი ნაჩვენებია მხოლოდ ჩამორჩენილ ჯაჭვზე. ფაქტობრივად, ისინი ლიდერობენ. თუმცა, აქ მხოლოდ ერთი პრაიმერია საჭირო თითო ჩანგალზე.
ვინაიდან დედის დნმ-ის ჯაჭვები ყოველთვის არ განსხვავდება ბოლოებიდან, მაგრამ ინიციალიზაციის წერტილებში, სინამდვილეში, არც ისე ბევრი ჩანგალი იქმნება, როგორც თვალები, ან ბუშტები.
თითოეულ ბუშტს შეიძლება ჰქონდეს ორი ჩანგალი, ანუ ჯაჭვები განსხვავდებიან ორი მიმართულებით. თუმცა მათ მხოლოდ ერთი შეუძლიათ. თუ, მიუხედავად ამისა, შეუსაბამობა ორმხრივია, მაშინ დნმ-ის ერთ ჯაჭვზე ინიციალიზაციის წერტილიდან სინთეზი წავა ორი მიმართულებით - წინ და უკან. ამ შემთხვევაში უწყვეტი სინთეზი შესრულდება ერთი მიმართულებით, ხოლო ოკაზაკის ფრაგმენტები მეორე მიმართულებით.
პროკარიოტული დნმ არ არის წრფივი, მაგრამ აქვს წრიული სტრუქტურა და მხოლოდ ერთი რეპლიკაციის საწყისი.
დიაგრამაზე, დნმ-ის მთავარი მოლეკულის ორი ჯაჭვი ნაჩვენებია წითლად და ლურჯად. სინთეზირებული ახალი ძაფები ნაჩვენებია წერტილოვანი ხაზებით.
პროკარიოტებში დნმ-ის თვითკოპირება უფრო სწრაფია, ვიდრე ევკარიოტებში. თუ ევკარიოტებში რედუპლიკაციის სიჩქარე წამში ასობით ნუკლეოტიდია, მაშინ პროკარიოტებში ის ათასს ან მეტს აღწევს.
რეპლიკაციის ფერმენტები
დნმ-ის რეპლიკაცია ხორციელდება ფერმენტების კომპლექსით, რომელსაც ე.წ საპასუხო. სულ 15-ზე მეტი რეპლიკაციის ფერმენტი და ცილაა, ყველაზე მნიშვნელოვანი ქვემოთ ჩამოთვლილია.
რეპლიკაციის მთავარი ფერმენტი უკვე აღნიშნულია დნმ პოლიმერაზა(ფაქტობრივად, არსებობს რამდენიმე განსხვავებული), რაც პირდაპირ ახორციელებს ჯაჭვის ზრდას. ეს არ არის ფერმენტის ერთადერთი ფუნქცია. პოლიმერაზას შეუძლია „შეამოწმოს“, რომელი ნუკლეოტიდი ცდილობს ბოლოს შეერთოს. თუ არ არის შესაფერისი, მაშინ ის წაშლის მას. სხვა სიტყვებით რომ ვთქვათ, დნმ-ის ნაწილობრივი შეკეთება, ანუ მისი რეპლიკაციის შეცდომების კორექტირება ხდება უკვე სინთეზის ეტაპზე.
ნუკლეოპლაზმაში (ან ციტოპლაზმაში ბაქტერიებში) ნაპოვნი ნუკლეოტიდები არსებობს ტრიფოსფატების სახით, ანუ ისინი არ არიან ნუკლეოტიდები, არამედ დეოქსინუკლეოზიდური ტრიფოსფატები (dATP, dTTP, dGTP, dCTP). ისინი ჰგავს ATP-ს, რომელსაც აქვს სამი ფოსფატის ნარჩენი, რომელთაგან ორი დაკავშირებულია მაკროერგიული ობლიგაციებით. როდესაც ეს ობლიგაციები იშლება, ბევრი ენერგია გამოიყოფა. ასევე, დეოქსინუკლეოზიდის ტრიფოსფატებში, ორი ბმა მაკროერგიულია. პოლიმერაზა გამოყოფს ბოლო ორ ფოსფატს და იყენებს გამოთავისუფლებულ ენერგიას დნმ-ის პოლიმერიზაციის რეაქციისთვის.
ფერმენტი ჰელიკაზაჰყოფს მატრიქსის დნმ-ის ძაფებს მათ შორის წყალბადის ბმების გაწყვეტით.
ვინაიდან დნმ-ის მოლეკულა ორმაგი სპირალია, ობლიგაციების გაწყვეტა კიდევ უფრო მეტ გადახვევას იწვევს. წარმოიდგინეთ თოკი ორი თოკისგან, რომლებიც ერთმანეთთან შედარებით გადაუგრიხეს და ერთ მხარეს ბოლოები აწიეთ მარჯვნივ, მეორე მარცხნივ. ნაქსოვი ნაწილი კიდევ უფრო დაგრეხილი გახდება, უფრო მჭიდრო იქნება.
ასეთი დაძაბულობის აღმოსაფხვრელად აუცილებელია, რომ ორმაგი სპირალი, რომელიც ჯერ კიდევ არ დაიშალა, სწრაფად ბრუნავდეს თავისი ღერძის გარშემო და "გადააყენოს" შედეგად მიღებული ზეგადახვევა. თუმცა, ეს ძალიან ენერგო ინტენსიურია. ამიტომ უჯრედებში განსხვავებული მექანიზმი რეალიზდება. ფერმენტი ტოპოიზომერაზაარღვევს ერთ-ერთ ძაფს, მეორეს უღელტეხილზე გადის და პირველს ისევ კერავს. ეს აღმოფხვრის წარმოქმნილ სუპერკოჭებს.
მატრიქსის დნმ-ის ძაფები, რომლებიც ჰელიკაზას მოქმედების შედეგად გაიფანტა, ცდილობენ ხელახლა დაუკავშირდნენ წყალბადურ ბმებს. ამის თავიდან ასაცილებლად, დნმ-ის დამაკავშირებელი ცილები. ისინი არ არიან ფერმენტები იმ გაგებით, რომ ისინი არ ახდენენ რეაქციების კატალიზებას. ასეთი ცილები მიმაგრებულია დნმ-ის ჯაჭვზე მთელ სიგრძეზე და არ იძლევა შაბლონის დნმ-ის დამატებითი ჯაჭვების დახურვის საშუალებას.
სინთეზირებულია პრაიმერები რნმ პრიმაზა. და ამოღებულია ეგზონუკლეაზა. პრაიმერის ამოღების შემდეგ, სხვა ტიპის პოლიმერაზა აყალიბებს "ხვრელს". თუმცა, ამ შემთხვევაში, დნმ-ის ცალკეული მონაკვეთები არ არის შეკერილი.
სინთეზირებული ჯაჭვის ცალკეული ნაწილები ჯვარედინი კავშირშია ისეთი რეპლიკაციის ფერმენტით, როგორიცაა დნმ ლიგაზა.